Exemplos de genes alélicos e não alélicos. Interação de genes alélicos
Genética humana com noções básicas de genética geral [Tutorial] Kurchanov Nikolay Anatolyevich
4.3. Interação genética
4.3. Interação genética
Muitos genes funcionam simultaneamente no corpo. Nos processos de implementação da informação genética numa característica, numerosos “pontos” de interação entre diferentes genes são possíveis ao nível das reações bioquímicas. Tais interações afetam inevitavelmente a formação do fenótipo.
Os genes alélicos determinam características alternativas, uma vez que se encontram em loci homólogos de cromossomos homólogos. Certas interações são possíveis entre os alelos de um heterozigoto, que fundamentam a manifestação da característica no fenótipo. São conhecidas três formas principais de interações interalélicas.
Dominância completa - em um heterozigoto, um alelo (dominante) suprime a manifestação do segundo alelo (recessivo).
Dominância incompleta - em um heterozigoto há uma manifestação intermediária das características dos alelos.
A codominância é a manifestação independente de alelos em um heterozigoto.
Em alguns casos, o mecanismo de interação dos alelos foi decifrado. Sua relação é melhor ilustrada por proteínas enzimáticas. Dados da genética bioquímica mostram que o fenômeno da dominância está frequentemente associado à atividade de uma enzima específica.
Menos claro é o fenômeno da heterose monogênica, quando um heterozigoto exibe uma característica mais fortemente do que um homozigoto para o alelo dominante. Heterose- o fenômeno da superioridade dos híbridos sobre ambas as formas parentais. O fenômeno da heterose tem sido intensamente estudado devido ao seu importante papel na seleção. Este fenômeno é muito complexo e ambíguo. O conceito de “superioridade” é ambíguo, uma vez que a heterose se manifesta nos níveis reprodutivo, somático e adaptativo. As manifestações em diferentes níveis podem ocorrer em direções opostas (por exemplo, uma “melhoria” nos indicadores somáticos pode ser acompanhada por uma “deterioração” nos adaptativos). Os próprios conceitos de “melhoria” e “deterioração” são ambíguos, pois não possuem critérios claros e sua aplicação é extremamente subjetiva. Existem várias teorias que explicam a natureza da heterose.
O fenômeno da heterose monogênica, quando é demonstrada a dependência de uma característica de um par alélico, é apenas um aspecto da teoria da heterose. Por sugestão de F. Dobzhansky, esse fenômeno foi denominado superdominância.É de grande importância para a teoria evolutiva porque demonstra a vantagem dos heterozigotos nas populações. No entanto, este conceito é bastante aplicável ao nível adaptativo, portanto, até que os mecanismos genéticos de sobredominância sejam esclarecidos, é prematuro considerá-lo como um tipo especial de interações inter-alélicas.
Nos organismos vivos, não apenas os alelos de um gene interagem frequentemente, mas também alelos de genes diferentes, proporcionando uma ampla variedade de opções de segregação. Existem três tipos principais de interação entre genes não alélicos.
Complementaridade – a interação de diferentes alelos dominantes provoca o aparecimento de uma nova característica.
Genes que controlam diferentes estágios de uma mesma via metabólica geralmente interagem de acordo com o tipo de complementaridade. Contudo, para alguns caracteres morfológicos o mecanismo bioquímico de implementação é desconhecido.
Epistasia - um gene suprime a expressão de outro gene não alélico.
Os genes que suprimem a ação de outros genes são chamados epistáticos (ou genes supressores). Existem duas opções possíveis para epistasia: epistasia dominante– o gene epistático é dominante em seu par alélico e epistasia recessiva– um gene epistático é recessivo em seu par alélico.
Polimerismo é a ação inequívoca de genes não alélicos. O polimerismo está associado ao controle da característica por vários genes não alélicos. O controle poligênico é muito difundido na genética. Os genes do polímero são geralmente designados pelas mesmas letras com um subscrito - A 1, A 2, A 3, etc.
A polimeria também ocorre em duas variedades. No polímero cumulativo a intensidade da característica é proporcional ao número de alelos dominantes entre os genes poliméricos, e quando polímero não cumulativo diferentes genes poliméricos se duplicam e, para que a característica se manifeste, a presença de um dos alelos dominantes é suficiente.
Numerosos casos de interação genética preenchem a maior parte de todos os livros sobre problemas genéticos. Num caso típico, ao cruzar diheterozigotos, a interação de genes resulta na formação de proporções muito diferentes de classes fenotípicas em gerações - 9:3:4; 9:7; 13:3; 12:3:1; 15:1 e outros. A análise genética mostra que são todas modificações da fórmula clássica de clivagem diíbrida mendeliana 9: 3: 3: 1. Resolver um grande número de problemas em genética é uma etapa necessária na preparação de estudantes que estudam genética.
A frase “interação genética” é um tanto arbitrária, uma vez que geralmente não são os próprios genes que interagem, mas seus produtos. Contudo, não podemos concordar com o termo “interação dos secadores de cabelo”, que não reflete com precisão o significado do fenômeno. Na minha opinião, é melhor deixar o termo tradicional “interação genética” (alélico e não alélico) na literatura educacional.
Este texto é um fragmento introdutório. Do livro Genética das Cores dos Cachorros por Robinson RoyInteração dos loci A e E A importância dessas duas séries de alelos será mostrada nesta seção. Ambas as séries controlam a distribuição dos pigmentos pretos e amarelos pela pelagem da maioria das raças de cães. Em interação, esses loci formam muitos
Do livro Microbiologia: notas de aula autor Tkachenko Ksenia Viktorovna2. Interação dos vírus com a célula hospedeira A interação ocorre em um único sistema biológico no nível genético. Existem quatro tipos de interação: 1) infecção viral produtiva (interação que resulta na reprodução do vírus, e
Do livro Formiga, Família, Colônia autor Zakharov Anatoly AlexandrovichINTERAÇÃO DE FORMIGAS EM UMA FAMÍLIA Uma família de formigas é uma associação de centenas, milhares e às vezes milhões de indivíduos, por meio de cujos esforços constroem um ninho, alimentam numerosos filhotes, protegem o formigueiro e sua área de alimentação das invasões de agressivos
Do livro Evolução autor Jenkins MortonDERIVA GENE Este conceito é às vezes chamado de “efeito Sewell-Wright”, em homenagem aos dois geneticistas populacionais que o propuseram. Depois que Mendel provou que os genes são as unidades da hereditariedade, e Hardy e Weinberg demonstraram o mecanismo de seu comportamento,
Do livro Raça Humana por Barnett AnthonyInteração entre hereditariedade e meio ambiente Às vezes as pessoas perguntam: o que é mais importante - hereditariedade ou meio ambiente? Esta pergunta não é tão fácil de responder. Se com isso queremos dizer o quê? tem o maior impacto, então, mesmo assim, deveríamos limitar-nos a casos especiais.
Do livro Fundamentos de Psicofisiologia autor Alexandrov Yuri7. INTERAÇÃO DOS SISTEMAS SENSORAIS A interação dos sistemas sensoriais ocorre nos níveis espinhal, reticular, talâmico e cortical. A integração de sinais na formação reticular é especialmente ampla. A integração de sinais de ordem superior ocorre no córtex cerebral. EM
Do livro Embriões, Genes e Evolução por Raff Rudolf AInteracção e integração Hoje em dia, os visitantes dos museus consideram como garantidos os esqueletos montados de enormes animais extintos em exposição. No entanto, nem sempre foi assim. Informações científicas sobre formas impressionantes como mamutes e
Do livro Teoria da Nutrição Adequada e Trofologia [tabelas no texto] autor Do livro Pare, quem lidera? [Biologia do comportamento de humanos e outros animais] autor Jukov. Dmitry AnatolyevichInteração dos sistemas endócrinos: comunicação direta, feedback, sinergismo, ação permissiva, antagonismo Como já mencionado, as glândulas endócrinas individuais e seus hormônios interagem entre si, garantindo a unidade funcional do corpo. Entre
Do livro Teoria da Nutrição Adequada e Trofologia [tabelas com fotos] autor Ugolev Alexander Mikhailovich9.7. Observações finais (interação em biocenoses) Consideremos as interações bioquímicas dos organismos vivos em condições naturais, que são extremamente difundidas e se tornaram objeto de uma ciência especial - a alelopatia (ver Capítulo 1). Exemplos de tais
Do livro Fenética [Evolução, população, característica] autor Yablokov Alexei VladimirovichInteração de forças evolutivas - o mecanismo de evolução O mecanismo desencadeador da evolução funciona como resultado da ação conjunta de fatores evolutivos dentro da população como uma unidade evolutiva. Cada população de cada espécie está sujeita a pressões de um tipo ou de outro.
Do livro Evolução [Ideias clássicas à luz de novas descobertas] autorDuplicação de genes GENES MULTIFUNCIONAIS - A BASE DAS INOVAÇÕES EVOLUCIONÁRIAS A ideia de que a duplicação de genes serve como a fonte mais importante de inovações evolutivas foi expressa na década de 1930 pelo notável biólogo John Haldane (Haldane, 1933). Hoje não há
Do livro Genes e o desenvolvimento do corpo autor Neyfakh Alexander Alexandrovich1. Promotores de genes Nesta seção falaremos brevemente sobre quais sequências de nucleotídeos adjacentes aos genes, e às vezes dentro de um gene, são responsáveis pelo processo de transcrição. Nos procariontes, essas regiões às quais a molécula de RNA polimerase se liga e de onde
Do livro Evolução Humana. Livro 1. Macacos, ossos e genes autor Markov Alexander VladimirovichMudanças na atividade genética A evolução dos animais em geral e dos primatas em particular ocorre não tanto devido a mudanças na estrutura dos genes codificadores de proteínas, mas sim devido a mudanças em sua atividade. Pequena mudança nos andares superiores organizados hierarquicamente
Do livro Genética Humana com Noções Básicas de Genética Geral [Tutorial] autor Kurchanov Nikolai Anatolievich4.4. Interação genótipo e ambiente A natureza da manifestação da ação dos genes é muito mais complexa do que nas opções descritas acima. Ao considerar o efeito dos genes e seus alelos, é necessário levar em consideração a influência do ambiente externo na manifestação dos traços, bem como o efeito modificador de outros
Do livro do autor7.1. Isolamento de genes Podem ser utilizadas várias maneiras de isolar genes. Cada um deles tem suas vantagens e desvantagens.A síntese química de genes, ou seja, a síntese de nucleotídeos com uma determinada sequência correspondente a um gene, foi realizada pela primeira vez em
A interação entre genes alélicos ocorre de três formas: dominância completa, dominância incompleta e manifestação independente (codominância).
Dominância completa - quando um alelo dominante suprime completamente a manifestação de um alelo recessivo, por exemplo, a cor amarela das ervilhas é dominante sobre a cor verde.
A dominância incompleta é observada quando um gene de um par de alelos não proporciona a formação de seu produto proteico suficiente para a manifestação normal da característica. Com esta forma de interação genética, todos os heterozigotos e homozigotos diferem significativamente uns dos outros no fenótipo. Um exemplo de divisão com dominância incompleta é a herança da cor das flores da Beleza Noturna.
Ao cruzar plantas com flores vermelhas (AA) e plantas com flores brancas (aa), os híbridos F1 apresentam flores rosa (Aa). Assim, ocorre dominância incompleta; em F2 há uma divisão 1:2:1 tanto no fenótipo quanto no genótipo.
Além da dominância completa e incompleta, são conhecidos casos de ausência de relações dominante-recessivas ou co-dominância. Quando codominante em organismos heterozigotos, cada um dos genes alélicos causa a formação de uma característica controlada por ele no fenótipo.
Um exemplo dessa forma de interação de alelos é a herança de grupos sanguíneos humanos segundo o sistema AB0, determinado pelo gene I. Existem três alelos desse gene Io, Ia, Ib, que determinam os antígenos dos grupos sanguíneos. A herança dos grupos sanguíneos também ilustra o fenômeno do alelismo múltiplo: nos pools genéticos das populações humanas, o gene I existe na forma de três alelos diferentes, que são combinados em indivíduos individuais apenas aos pares. Antes deste exemplo, falamos sobre genes que existem apenas em duas formas alélicas diferentes. No entanto, muitos genes consistem em centenas de pares de nucleotídeos, de modo que mutações podem ocorrer em muitas regiões do gene e dar origem a muitas formas alélicas diferentes. Como cada um dos cromossomos homólogos possui um gene alélico, então, é claro, um organismo diplóide não possui mais do que dois de uma série de alelos no pool genético da população.
30. Interações genéticas não alélicas
Genes não alélicos- são genes localizados em diferentes partes dos cromossomos e que codificam diferentes proteínas. Genes não-alélicos também podem interagir uns com os outros.
Nesse caso, ou um gene determina o desenvolvimento de vários traços ou, inversamente, um traço se manifesta sob a influência de uma combinação de vários genes. Existem três formas e interações de genes não alélicos:
complementaridade;
polimerismo.
Complementar A ação genética (adicional) é um tipo de interação de genes não alélicos, cujos alelos dominantes, quando combinados em um genótipo, determinam uma nova manifestação fenotípica de características. Neste caso, a divisão dos híbridos F2 de acordo com o fenótipo pode ocorrer nas proporções 9:6:1, 9:3:4, 9:7, às vezes 9:3:3:1. Um exemplo de complementaridade é a herança do formato do fruto da abóbora. A presença dos genes dominantes A ou B no genótipo determina o formato esférico do fruto e o formato alongado dos genes recessivos. Se o genótipo contiver os genes dominantes A e B, o formato do fruto será discóide. Ao cruzar linhagens puras com variedades que possuem formato de fruto esférico, na primeira geração híbrida F1 todos os frutos terão formato de disco, e na geração F2 haverá uma divisão no fenótipo: de cada 16 plantas, 9 terá frutos em forma de disco, 6 - esféricos e 1 - alongado.
Epistasia- interação de genes não alélicos, em que um deles é suprimido pelo outro. O gene supressor é denominado epistático, o gene suprimido é denominado hipostático. Se um gene epistático não tem manifestação fenotípica própria, ele é chamado de inibidor e é designado pela letra I. A interação epistática de genes não alélicos pode ser dominante e recessiva. Na epistasia dominante, a manifestação do gene hipostático (B, b) é suprimida pelo gene epistático dominante (I > B, b). A clivagem do fenótipo durante a epistasia dominante pode ocorrer na proporção 12:3:1, 13:3, 7:6:3. Epistasia recessiva é a supressão por um alelo recessivo de um gene epistático de alelos de um gene hipostático (i > B, b). A clivagem fenotípica pode ocorrer na proporção 9:3:4, 9:7, 13:3.
Polimerismo- interação de múltiplos genes não alélicos que influenciam claramente o desenvolvimento da mesma característica; O grau de manifestação de uma característica depende do número de genes. Os genes do polímero são designados pelas mesmas letras e os alelos do mesmo locus têm o mesmo subscrito.
A interação polimérica de genes não alélicos pode ser cumulativa e não cumulativa. Com a polimerização cumulativa (acumulativa), o grau de manifestação da característica depende da ação somativa dos genes. Quanto mais alelos genéticos dominantes, mais pronunciada é uma característica específica. A clivagem de F2 no fenótipo ocorre na proporção 1:4:6:4:1.
Com a polimerização não cumulativa, a característica se manifesta na presença de pelo menos um dos alelos dominantes dos genes do polímero. O número de alelos dominantes não afeta o grau de expressão da característica. A segregação fenotípica ocorre numa proporção de 15:1.
Exemplo: cor da pele em humanos, que depende de quatro genes.
Genes alélicos são genes localizados em regiões idênticas de cromossomos homólogos e controlando o desenvolvimento de variações de uma característica.
Os genes não-alélicos estão localizados em diferentes partes dos cromossomos homólogos e controlam o desenvolvimento de várias características.
O conceito de ação genética.
Um gene é uma seção de uma molécula de DNA ou RNA que codifica uma sequência de nucleotídeos em tRNA e rRNA ou uma sequência de aminoácidos em um polipeptídeo.
Características da ação genética:
O gene é discreto
O gene é específico - cada gene é responsável pela síntese de uma substância estritamente específica
O gene atua gradualmente
Efeito pleiotrópico - 1 gene atua na alteração ou manifestação de diversos sinais (Placa 1910) fenilcetonúria, síndrome de Marfan
Ação do polímero – vários genes são necessários para a expressividade de uma característica (1908 Nilsson-Ehle)
Os genes interagem entre si através de produtos proteicos determinados por eles
A expressão gênica é influenciada por fatores ambientais
Liste os tipos de interações entre genes alélicos e não alélicos.
Entre alelos:
Dominação Completa
Dominância incompleta
Codominância
Excesso de domínio
Entre não alélicos: (uma característica ou propriedades são determinadas por dois ou mais genes não alélicos que interagem entre si. Embora aqui a interação seja condicional, porque não são os genes que interagem, mas os produtos controlados por eles. Em neste caso, há um desvio das leis mendeleevianas de segregação).
Polimerismo
Complementaridade
A essência do domínio completo. Exemplos.
A dominância completa é um tipo de interação de genes alélicos em que o gene dominante (A) suprime completamente a ação do gene recessivo (a) (sardas)
Dominância incompleta. Exemplos.
Dominância incompleta é um tipo de interação de genes alélicos em que o alelo dominante não suprime completamente a ação do alelo recessivo, formando um traço com grau intermediário de degeneração (cor dos olhos, formato do cabelo)
A superdominância como base da heterose. Exemplos.
A sobredominância é um tipo de interação de genes alélicos em que um gene em estado heterozigoto tem uma manifestação fenotípica de uma característica maior do que um gene homozigoto.
Anemia falciforme. A – hemoglobinaA, e – hemoglobinaS. AA – glóbulos vermelhos 100% normais, mais suscetíveis à malária; aa – 100% mutado (morre), Aa – 50% mutado, praticamente não suscetível à malária porque já estou maravilhado
Codominância e sua essência. Exemplos.
Codominância é um tipo de interação de genes alélicos em que vários alelos de um gene estão envolvidos na determinação de uma característica e uma nova característica é formada. Um gene alélico complementa a ação de outro gene alélico, a nova característica difere das parentais (grupo sanguíneo ABO).
O fenômeno da manifestação independente de ambos os alelos no fenótipo de um heterozigoto, ou seja, a ausência de relações dominante-recessivas entre os alelos. O exemplo mais famoso é a interação de alelos que determinam o quarto grupo sanguíneo humano (AB). É conhecida uma série múltipla, composta por três alelos do gene I, que determina o sinal do grupo sanguíneo de uma pessoa. O gene I é responsável pela síntese de enzimas que ligam certos polissacarídeos a proteínas localizadas na superfície dos glóbulos vermelhos. (Esses polissacarídeos na superfície dos glóbulos vermelhos determinam a especificidade dos grupos sanguíneos.) Os alelos 1 A e 1 B codificam duas enzimas diferentes; o alelo 1° não codifica nada. Neste caso, o alelo 1° é recessivo em relação a 1 A e I B, e não há relação dominante-recessiva entre os dois últimos. Pessoas com o quarto grupo sanguíneo carregam dois alelos em seu genótipo: 1 A e 1 B. Como não existe uma relação dominante-recessiva entre esses dois alelos, ambas as enzimas são sintetizadas no corpo dessas pessoas e o fenótipo correspondente é formado - o quarto grupo sanguíneo.
Domínio completo. Usando o exemplo dos experimentos de G. Mendel com ervilhas, fica claro que um gene pode suprimir completamente a manifestação de outro gene alélico.
Dominância incompleta. Com dominância incompleta, o traço também aparece na primeira geração intermediária entre as formas parentais. Se você acasalar ovelhas Karakul de orelhas compridas com ovelhas sem orelhas, então na F1 todos os descendentes terão orelhas curtas. A divisão em F2 por fenótipo e genótipo coincide (1:2:1).
Codominância. Ambos os genes alélicos aparecem no fenótipo, não revelando dominância nem recessividade entre si. Em humanos, são conhecidos os sistemas sanguíneos ABO, que determinam antígenos eritrocitários. Os alelos A e B interagem de acordo com o tipo de codominância.
De 1940 a 1970, 12 sistemas genéticos de grupos sanguíneos foram descobertos em bovinos. Eles distinguem mais de 100 antígenos sanguíneos, definindo 369 fenogrupos, que podem equivaler a cerca de dois trilhões de combinações diferentes – tipos sorológicos. Fenogrupo- um grupo de antígenos (às vezes um antígeno), que são a expressão fenotípica de um conjunto de genes individuais de um locus. Os grupos sanguíneos, que se baseiam nas características individuais das propriedades antigênicas dos glóbulos vermelhos, são hereditários e não mudam ao longo da vida do animal. Eles são herdados de forma única ou complexa e, portanto, podem servir como um modelo genético conveniente para resolver muitas questões teóricas e práticas de seleção, uma vez que a grande maioria dos fatores alélicos eritrocitários conhecidos são herdados de acordo com o tipo de codominância.
Excesso de domínio. Caracteriza-se pelo fato de que em indivíduos heterozigotos o traço é mais pronunciado do que em indivíduos homozigotos dominantes, ou seja, o híbrido é superior a ambos os pais no desenvolvimento do traço. Este fenômeno é denominado heterozas (Aa>AA e aa). Um exemplo claro de excesso de domínio é a anemia falciforme humana. Os homozigotos SS têm alguns de seus glóbulos vermelhos em forma de foice e ponta de flecha, e o transporte de oxigênio pelos glóbulos vermelhos é prejudicado. As pessoas sofrem de anemia aguda e geralmente morrem precocemente. Os heterozigotos Ss são resistentes à malária tropical, os homozigotos ss desenvolvem malária tropical.
Alelismo múltiplo. Consideramos principalmente a interação de dois genes alélicos, cada um dos quais pode mudar como resultado de uma mutação e surgir um novo gene alélico. Este fenômeno é denominado "alelismo múltiplo". Em humanos, um exemplo de alelismo múltiplo é o sistema sanguíneo ABO, que possui 3 genes alélicos (A, B, O) e, consequentemente, 6 genótipos e 4 fenótipos.
Os seguintes alelos foram identificados em coelhos que afetam a cor do cabelo: C - preto, cсh - chinchila, ch - Himalaia, ca - albino. Os alelos são apresentados em ordem decrescente de dominância (C>cch>ch>ca). O alelo C é dominante sobre todos os subsequentes, ca é recessivo para todos os anteriores (Fig. 2.5).
Efeito pleiotrópico do gene. Um gene pode influenciar o desenvolvimento de duas ou mais características, ou seja, são observados múltiplos efeitos do gene. Este tipo de interação foi descoberto durante a criação de ovelhas Karakul cinzentas (Shirazi) e pretas (Arabi). Cordeiros homozigotos para o alelo dominante do gene C, que determina a cor cinza (Shirazi). ao passarem para o pasto morrem devido ao subdesenvolvimento do sistema nervoso parassimpático do rúmen. Isso se deve à ação de dois genes dominantes, que têm duplo efeito (pleiotrópico): causam coloração acinzentada e, no estado homozigoto, defeito no sistema nervoso.
O acadêmico D. K. Belyaev e seus colegas descobriram que em visons algumas mutações associadas a mudanças na cor do cabelo são recessivas e, devido ao efeito pleiotrópico, a fertilidade e a viabilidade dos animais são reduzidas.
Genes letais e semilegais. Os genes causam mudanças fisiológicas complexas durante o desenvolvimento de um organismo. Eles podem reduzir a vitalidade do corpo, até ao ponto da morte.
Um dos motivos que altera a divisão na proporção de 3:1. é a viabilidade diferente dos homozigotos em F2. No século 20 fatos foram descobertos quando ocorreu uma divisão do fenótipo 2:1. Assim, ao estudar a herança da cor em camundongos amarelos e pretos, foi observada uma divisão na cor em 2 partes de camundongos amarelos e 1 parte de camundongos pretos. Neste caso, alguns dos ratos amarelos morreram durante o período embrionário. O mesmo gene no estado heterozigoto dominante causou a coloração amarela dos camundongos e no estado homozigoto mostrou seu efeito letal. O gene da cor preta é alélico e recessivo.
Na década de 30 do século XX. Uma nova coloração foi descoberta na raposa preta prateada, chamada platina. Quando raposas platinadas foram cruzadas entre si, os indivíduos apresentavam coloração platina e preta prateada na proporção de 2:1. Ao realizar um cruzamento analítico, ficou claro que o gene da cor platina é dominante. Foi sugerido que raposas platinadas homozigotas (AA) morrem durante o período embrionário. Autópsias de mulheres grávidas confirmaram esta suposição.
Os genes letais recessivos atuam no estado homozigoto, e os dominantes também atuam no estado heterozigoto. Se um gene não causa a morte do organismo, mas reduz significativamente a viabilidade, então é denominado semiletal.
Genes letais e semilegais são geralmente recessivos. Genes letais causam a morte do organismo no período embrionário ou pós-embrionário. Por exemplo, genes legais recessivos causam a ausência de pêlo em bezerros recém-nascidos, coelhos e animais de outras espécies. Logo após o nascimento, esses animais morrem.
Genes semilegais podem causar deformidades em animais: encurtamento do esqueleto axial; deformação da cabeça e membros; violação das funções fisiológicas do corpo. Os animais reprodutores com genes letais e semiletais devem ser abatidos. É especialmente necessário verificar os reprodutores dos quais um grande número de descendentes é obtido através de inseminação artificial.
Os genes legais podem espalhar-se livremente de um país para outro. Assim, o touro reprodutor da raça holandesa Prince Adolf, portador de uma anomalia genética, foi trazido da Holanda para a Suécia em 1902. 26 anos depois, soube-se que o touro é portador do gene recessivo letal para bezerros sem pêlo. Suas filhas, filhos e netos foram amplamente utilizados para fins de reprodução e, em um tempo relativamente curto, muitos dos animais foram parentes do Príncipe Adolfo. Em alguns rebanhos, cerca de 6% dos bezerros nasceram sem pelos e morreram poucos minutos após o nascimento.
Recentemente, surgiram relatos na literatura nacional e estrangeira sobre anomalias genéticas (defeitos) que causam enormes danos econômicos à pecuária (P. D. Smith, L. F. Novikova). Assim, por exemplo, defeitos BLAD (Deficiência de Adesão de Leucócitos Bovinos), ou adesão (aglomeração) de leucócitos em bovinos e CVM (Malformação Vertebral Complexa), ou deformidade complexa da coluna vertebral, têm um tipo de herança recessiva e não são expressos no estado heterozigoto (Fig. 2.6) . Esses defeitos genéticos se espalharam na linha de gado Holstein Montvik Chieftain 95679, que é amplamente utilizado na Federação Russa. Um dos touros mais famosos desta linha, Osbordale Ivanhoe 1189870, era portador do defeito genético B LAD. e seu neto Carlin-M Ivanhoe Bell 1667366 é portador de defeitos genéticos BLAD e CVM.
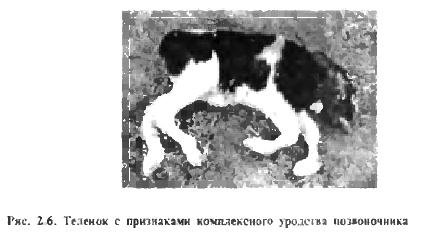
Em média, 2 a 5% dos animais recém-nascidos de muitas espécies apresentam anomalias hereditárias. Genes letais e semilegais são uma das causas de mortalidade embrionária e anomalias congênitas. Várias dezenas desses genes foram identificados para cada tipo de animal de criação e foi fornecida uma classificação internacional de anomalias. A natureza hereditária dos nados-mortos, abortos espontâneos e deformidades é evidenciada pelo facto de a sua percentagem aumentar significativamente com a endogamia.
As anomalias hereditárias mais comuns em bovinos são: tipo pug; contratura muscular; ausência de membros anteriores e posteriores; hidropisia do feto e do cérebro; defeitos da pele: falta de pelos; encurtamento da mandíbula; hérnia umbilical, etc.
Resumindo os dados de diversos autores, podemos dizer que no gado existem mais de 40 letalidades homozigóticas, deformidades e anomalias cariotípicas - casamentos hereditários. Foram identificados 23 defeitos congênitos que afetam a função reprodutiva e a fertilidade das vacas.
33 anomalias genéticas foram descritas em porcos. Os mais comuns são: hérnia cerebral; paralisia dos membros posteriores; ausência de ânus; fenda palatina; pernas grossas; curvatura e rigidez dos membros; hidropisia do cérebro; hemofilia.
Em ovinos, os defeitos craniofaciais mais pronunciados são: encurtamento ou ausência da mandíbula; perda de cabelo, calvície; nanismo; fenda palatina, etc. Um total de 43 anomalias.
A Lista Internacional de Defeitos Legais inclui 45 anomalias em galinhas; 6 - em perus; 3 - em patos; Das anomalias hereditárias, 10 ocorrem em cavalos.
A origem hereditária da anomalia é indicada pelos seguintes fatos: a correspondência da anomalia detectada com a descrição do defeito genético, o aparecimento da anomalia por endogamia, a manifestação de anomalias semelhantes em parentes laterais (irmãos e meio -irmãos), parentes da mãe ou do pai.
Todos os casos de manifestação de anomalias hereditárias devem ser registrados nas fichas dos reprodutores para que futuras análises genéticas possam ser realizadas. Nesta situação, o pedigree e os registros objetivos de melhoramento desempenham um papel importante. A coleta de dados para avaliar o bem-estar genético dos animais reprodutores e a presença de genes letais e semilegais deve ser realizada em criadouros e estações de inseminação artificial de fazendas. animais.
Interação de genes alélicos e seus tipos
Ao estudar as leis de G. Mendel, já conhecemos os principais tipos de interação dos genes alélicos. Com base no material previamente estudado, preencha a tabela.
Mesa. Tipos de interação entre genes de um par alélico
Tipo de interação |
A natureza da interação em |
Fórmulas para clivagem fenotípica em F 2 e em cruzamentos de teste |
Exemplos |
| Dominação Completa | Um alelo (dominante) suprime completamente o efeito de outro alelo (recessivo) |
A alta estatura das plantas domina o nanismo; cabelos cacheados - sobre cabelos lisos; olhos castanhos - acima dos azuis, etc. |
|
| Dominância incompleta | O efeito de cada alelo não é totalmente realizado, como resultado do aparecimento de uma característica intermediária |
Colorir flores de beleza noturna; anemia falciforme; cistonúria, etc. |
|
| Codominância | Participação de ambos os alelos na determinação da característica |
Grupo sanguíneo IV (os eritrócitos deste grupo sanguíneo possuem aglutinogênios do tipo A e digite EM) |
Interação de genes não-alélicos
Com base nas leis de G. Mendel, podemos chegar à conclusão de que existe uma conexão bastante forte entre um gene e uma característica, que o genótipo é composto pela soma de genes que atuam independentemente e o fenótipo é uma combinação mecânica de traços individuais. No entanto, conexões diretas e inequívocas entre um gene e uma característica são a exceção e não a regra. O desenvolvimento de uma característica em um organismo geralmente está sob o controle de muitos genes, e a característica é o resultado da interação de genes não alélicos.
O caso mais simples de interação de genes não alélicos é quando uma característica é controlada por dois pares de alelos. Vejamos alguns exemplos dessa interação.
Interação genética complementar
O fenômeno da interação gênica, em que a presença de dois genes não alélicos é necessária para a manifestação de uma característica, é denominado complementaridade(de lat. complemento- adição), e os genes necessários para a manifestação da característica são complementar, ou adicional.
Consideremos várias opções para interação genética complementar.
Alelos do segundo gene aparecem apenas na presença de um alelo dominante do geneA. A coloração dos ratos depende, no caso mais simples, de dois genes. Ratos com genótipo ahh carecem de pigmento e são de cor branca. Na presença de um alelo dominante A o pigmento é produzido e o mouse fica de alguma forma colorido. A cor específica é determinada pelo segundo gene. Seu alelo dominante COM causa a cor cinza do mouse e o alelo recessivo Com- cor preta. Assim, se considerarmos variantes homozigotas, o genótipo dos camundongos cinzentos é AASS, preto - AAAss, branco - aaSS ou burro. Ao cruzar um rato cinza AASS com branco burro na primeira geração tudo acontece segundo Mendel: todos os híbridos são cinza (são heterozigotos com o genótipo AaSs). Em F2, como é fácil de verificar, temos 9/16 ratos cinzentos, 3/16 pretos e 4/16 brancos.

Para que uma característica se manifeste, os alelos dominantes de dois genes diferentes devem estar presentes no genótipo. Um exemplo dessa interação genética complementar é a herança do formato dos frutos nas abóboras. Na presença de ambos os alelos dominantes, os frutos apresentam formato de disco, na presença de um (qualquer!) alelo dominante - esférico, e na ausência de alelos dominantes - alongados.

Para que uma característica se manifeste, os alelos dominantes de dois genes diferentes devem estar presentes no genótipo, mas cada alelo dominante, em combinação com os alelos recessivos do outro par, tem uma manifestação fenotípica independente. Por exemplo, em galinhas, a crista em forma de ervilha é determinada por um gene dominante, enquanto a crista em forma de rosa é determinada por outro gene não alélico, mas também dominante. Quando esses genes estão no mesmo genótipo, desenvolve-se uma crista em forma de noz. Se o organismo for homozigoto para ambos os genes recessivos, desenvolve-se um pente simples em forma de folha. Ao cruzar diíbridos (todos com favo em forma de noz), na segunda geração ocorre uma divisão na proporção de 9: 3: 3: 1. Mas aqui é impossível encontrar segregação independente de cada alelo na proporção de 3:1, pois em todos os casos de coincidência no genótipo de ambos os genes dominantes, seu efeito direto não é detectado.
Assim, falamos de interação complementar de genes não alélicos no caso em que dois genes não alélicos dão uma nova característica, ou seja, ocorre uma nova formação.
Interação genética epistática
A interação de genes, na qual um gene suprime a ação de outro, não alélico ao primeiro, é chamada epistasia(do grego epi– acima e estase- stand), e genes que suprimem a ação de outros genes são chamados genes supressores, ou genes inibidores(de lat. inibir– segurar), ou genes epistáticos. O gene que está sendo suprimido é chamado hipostático.
Existem epistasia dominante e recessiva.
Epistasia dominante associado a um gene inibidor dominante. Por exemplo, gene EU determina a cor branca dos frutos da abóbora; se presente, o efeito do gene EM não aparece. Com genótipo iiBB ou iiВb Os frutos da abóbora são amarelos. Finalmente, se ambos os genes forem representados por alelos recessivos, o fruto da abóbora é verde. O mesmo tipo pode ser usado para herdar a cor da pelagem de cães, ovelhas e outros animais.
Epistasia recessivaé detectado quando a expressão de um gene é suprimida por alelos recessivos de outro gene. Este tipo de epistasia é ilustrado pela herança da cor da pelagem em ratos domésticos. A cor da cutia (cinza avermelhado) é determinada pelo gene dominante A, seu alelo recessivo A dá uma cor preta. Gene de um par não alélico EM promove a manifestação da cor, e o gene bé um supressor e suprime o efeito do alelo dominante A e recessivo A. Ratos com genótipo Abb indistinguível no fenótipo de indivíduos com o genótipo aabb- tudo branco.
Interação genética polimérica
Polimerismo– um tipo de interação de genes não alélicos em que vários pares de genes não alélicos atuam no desenvolvimento de uma característica. Tais genes são chamados polímero(do grego política– muito e meros- Papel). Eles são denotados pela mesma letra, mas com índices diferentes, que indicam o número de pares alélicos no genótipo que determinam o desenvolvimento de uma determinada característica ( Um 1 Um 1 Um 2 Um 2).
Existem duas variantes de polimerização: com efeito de soma de genes e sem reforço de genes entre si.
O efeito cumulativo dos genes poliméricos. Nesse caso, o grau de manifestação da característica depende do número de alelos dominantes no genótipo do indivíduo. Assim, a cor vermelha dos grãos de trigo é determinada por dois ou mais pares de genes. Cada um dos genes dominantes desses alelos determina a cor vermelha, enquanto os genes recessivos determinam a cor branca dos grãos. Um alelo dominante produz grãos que não são muito coloridos. Se dois alelos dominantes estiverem presentes no genótipo, a intensidade da cor aumenta. Somente quando o organismo é homozigoto para todos os pares de genes recessivos os grãos não são coloridos. Assim, ao cruzar diíbridos, ocorre uma divisão na proporção de 15 coloridos para um branco. Mas de cada 15 pintadas, uma terá uma cor vermelha intensa, porque... contém quatro alelos dominantes, quatro terão uma cor ligeiramente mais clara, porque contêm três alelos dominantes e um alelo recessivo nos genótipos, seis são ainda mais leves com dois alelos dominantes e dois recessivos nos genótipos, quatro são ainda mais leves, porque têm apenas um gene dominante e três genes recessivos, ou seja, a verdadeira divisão será 1: 4: 6: 4: 1.
De acordo com esta versão do polímero, a cor da pele, a altura e o peso são herdados dos humanos. Um mecanismo semelhante de herança é característico de muitas características quantitativas, incluindo características economicamente valiosas: teor de açúcar nas raízes de beterraba, teor de vitaminas em frutas e vegetais, comprimento da espiga de cereais, comprimento da espiga de milho, fertilidade animal, produção de leite do gado , produção de ovos de galinhas, etc.
Os genes do polímero não se aprimoram. Neste caso, a divisão em F 2 será 15:1. Assim, os frutos da bolsa do pastor podem ser triangulares (dominantes) e ovais (recessivos). A característica é controlada por dois pares de genes poliméricos. Se o genótipo de uma planta tiver pelo menos um alelo dominante do primeiro ou segundo par de genes poliméricos, então o formato do fruto será triangular ( Um 1 Um 2; Um 1 um 2; um 1 A 2 ). Somente aquelas plantas que não possuem um único alelo dominante em seu genótipo terão fruto com formato oval - um 1 um 1 um 2 um 2.
Assim, o acúmulo de determinados alelos no genótipo pode levar a alterações na expressão das características.
Ação genética múltipla
 Freqüentemente, um gene afeta não uma, mas várias características de um organismo. O fenômeno no qual um gene pode influenciar a formação de diversas características de um organismo é denominado pleiotropia(do grego pleon– mais numerosos e tropos- vez).
Freqüentemente, um gene afeta não uma, mas várias características de um organismo. O fenômeno no qual um gene pode influenciar a formação de diversas características de um organismo é denominado pleiotropia(do grego pleon– mais numerosos e tropos- vez).
A existência deste fenómeno não contradiz de forma alguma o conceito clássico de “um gene – uma proteína – uma característica”. De forma simplificada, a influência de um gene em várias características ao mesmo tempo pode ser representada da seguinte forma. A partir da leitura das informações de um gene, forma-se uma determinada proteína, que pode então participar de diversos processos que ocorrem no organismo, exercendo assim múltiplos efeitos. Por exemplo, nas ervilhas, a cor marrom do tegumento da semente e o desenvolvimento do pigmento em outras partes da planta e a cor das flores dependem de um gene; na Drosophila, o gene que causa a cor branca dos olhos afeta o clareamento da cor do corpo e altera alguns órgãos internos; Nos humanos, o gene responsável pelos cabelos ruivos também determina a cor da pele mais clara e o aparecimento de sardas. A síndrome de Marfan é causada por um gene pleiotrópico autossômico dominante e se manifesta por alto crescimento, alongamento dos ossos dos dedos das mãos e dos pés (dedos de aranha), hipermobilidade articular, subluxação do cristalino e doenças cardíacas.
Essa ação múltipla é típica da maioria dos genes. No entanto, não se deve imaginar que um gene pleiotrópico influencia igualmente cada característica. A grande maioria dos genes com vários graus de pleiotropia são caracterizados por um efeito mais forte sobre uma característica e um efeito muito mais fraco sobre outra.
Genótipo como um sistema integral
O fato da divisão dos híbridos na prole permite afirmar que o genótipo é composto por elementos individuais - genes que podem ser herdados de forma independente ( discrição do genótipo). Ao mesmo tempo, o genótipo não pode ser considerado uma simples soma mecânica de genes individuais. Um genótipo é um sistema de genes em interação. Mais precisamente, não são os próprios genes (seções de DNA) que interagem, mas os produtos genéticos formados a partir deles (RNA e depois proteínas). Portanto, em alguns casos, a ação de diferentes genes é relativamente independente, mas, via de regra, a manifestação de uma característica é resultado da interação de produtos de diferentes genes.
O genótipo de qualquer organismo é um sistema complexo e integral de genes em interação. Essa integridade do genótipo surgiu historicamente no processo de evolução da espécie. Como resultado das mutações, novos genes aparecem constantemente, novos cromossomos e até novos genomas são formados. Os genes recém-surgidos podem interagir imediatamente com os genes existentes ou modificar a natureza do trabalho destes últimos, mesmo sendo recessivos, ou seja, sem se manifestarem.
Conseqüentemente, em cada espécie de plantas e animais o genótipo se manifesta como um sistema integral que se desenvolveu historicamente em um determinado momento.
II. Consolidação de conhecimento
Resumindo a conversa enquanto aprende novo material.
III. Trabalho de casa
Estude o parágrafo do livro didático (relação entre genes e características, tipos de interação de genes alélicos e não alélicos, pleiotropia, genótipo como sistema integral).
Resolva os problemas no final do parágrafo do livro didático.