Oddělení červených řas – rhodophyta. Oddělení červených řas – rhodophyta Význam pro člověka
Biologickým významem výskytu květenství je zvyšující se pravděpodobnost opylení květů entomofilních i anemofilních rostlin. Není pochyb o tom, že hmyz navštíví mnohem více květin za jednotku času, pokud jsou shromážděny v květenstvích. Kromě toho jsou květy shromážděné v květenstvích nápadnější mezi zelenými listy, spíše než jednotlivé květy. Mnoho povislých květenství se vlivem pohybu vzduchu snadno kymácí, čímž usnadňuje šíření pylu.
Květenství jsou charakteristická pro převážnou většinu rostlinných květů. Květenství jsou obvykle seskupena v blízkosti vrcholu rostliny na koncích větví, ale někdy, zejména u tropických stromů, se objevují na kmenech a silných větvích. Tento jev je znám jako caulifloria. Příkladem je čokoládový strom. Předpokládá se, že v podmínkách tropického lesa květák zpřístupňuje květy opylujícímu hmyzu. Dalším příkladem květáku je luštěnina Cercis rohovník, široce pěstovaná na Krymu a na Kavkaze.
Květenství má hlavní osu neboli osu květenství a postranní osy, které mohou být v různé míře rozvětvené nebo nevětvené. Jejich koncové větve – pedicely – nesou květy.
U jednoduchý Boční osy květenství nejsou rozvětvené a jsou pedicely. Květenství, ve kterých se rozvětvují postranní osy, se nazývají komplex. Složité květenství může mít boční osy prvního, druhého a následujících řádů.
Hlavní a všechny koncové boční osy mohou končit vrcholovými květy, v důsledku čehož je jejich růst omezený. Taková květenství se nazývají uzavřená ( sympodiální), nebo některé. V uzavřených květenstvích se vrcholové květy otevírají většinou dříve než podložní postranní, a proto se jim říká vrcholové květy.
V otevřených květenstvích je růst hlavní osy neomezený (tj. otevřený) a květy jsou umístěny na straně morfologického hrotu kvetoucího výhonku. Totéž může platit pro boční osy. Taková květenství se nazývají otevřená ( monopolní), nebo nedefinováno. V otevřených květenstvích kvetou květy postupně zdola nahoru, proto se jim říká boční kvetení.
2. Monopodiální, nebo botryodnye květenství.
Monopodiální (otevřená nebo postranně květovaná) květenství mají hlavní osu, která neustále roste, a postranní osy nesou květy. V takovém květenství vykvétají nejprve spodní květy a poté postupně horní. Existují jednoduchá a složitá botryoidní květenství
Jednoduchá botryoidní květenství
Štětec - na hlavní ose nebo ose prvního řádu jsou střídavě umístěny pedicely, které nesou drobné květy (konvalinka, třešeň ptačí, náprstník);
Jehněda je hlavní osou květenství, na rozdíl od hroznu visí dolů (bříza);
Třeň – na hlavní protáhlé ose jsou květy přisedlé (jitrocel);
Klas - hlavní osa květenství je velmi masitá, uspořádání květů je jako u klasu (kukuřice);
Sh i t o k - květy jsou umístěny ve stejné rovině, spodní květy mají delší stopky než horní (hruška, jablko, švestka);
Deštník - stopky jednotlivých květů jsou stejně dlouhé a vybíhají z horní části osy jako paprsky deštníku (cibule, třešeň);
Košík - četné přisedlé květy jsou umístěny na zkrácené a talířkovitě ztluštělé ose. Venku je obal z listenových listů (slunečnice, heřmánek);
Hlava – na zkrácené a rozšířené hlavní ose sedí blízko sebe květy (jetel, palina).
Složitá botryoidní květenství
Vyznačují se tím, že květy jsou umístěny na osách 2., 3. atd. řádu.
1. Panicle – na bočních osách jsou jednoduché štětce (lila, hrozny).
2. Složitý klásek – jednoduché klásky (pšenice, žito, ječmen) jsou umístěny na hlavní ose.
3. Složený deštník – na bočních osách jsou jednoduché deštníky. Jednoduché deštníky mají obvykle své zákrovy (soukromé zákrovy) a listeny na bázi deštníku tvoří společný zákrov (kopr, mrkev, anýz).
4. Scutellum komplexní (korymbózní lata) – na postranních osách jsou drobná květenství-koryty (jeřabina).
V některých případech jsou pozorována smíšená květenství. Například oves má latové květenství, ale místo květů jsou malé klásky, takže se mísí dvě květenství: lata a klas.
3. Sympodiální nebo cymoidní květenství. Morfologie ovoce.
Sympodiální (cymoidní) jsou ta květenství, u kterých je stopka (hlavní osa) zakončena květem a v jejím růstu pokračují horní postranní výhony, případně postranní výhony, které zase pokračují v růstu stejným způsobem. Cymoidní květenství zahrnují následující:
1. Vidlička, neboli dicházie. Hlavní osa končí květem, růst stopky pokračuje protilehlými postranními poupaty níže, tvořícími květy atd. (karafiát, koukol, mydlice).
2. Pleiochasium neboli falešný deštník – růst stopky končí květem a pokračuje vřetenově pod postranními pupeny, které tvoří květy (euphorbia).
3. Kudrnatka - růst stopky končí květem a pokračuje postranním pupenem tvořícím květ a tak dále opakovaně, přičemž všechny květy směřují jedním směrem (brambora, pomněnka, kostival).
Morfologie ovoce.
Po oplození se stěny pestíkového vaječníku vyvinou v oplodí (perikarp). Oplodí je stěna plodu obklopující semeno, která se tvoří současně s oplodím. Oplodí se obvykle skládá ze tří vrstev:
1) externí ( exokarp), někdy pokryté různými výrůstky (perutýn javorový, přívěsy lopuchu);
2) průměr ( mezokarp), který tvoří dužinu šťavnatého ovoce, obsahující hodně cukru (švestka, třešeň) nebo oleje (olivový);
3) vnitřní ( endokarp),často se mění ve vrstvu šťavnatého ovoce
kamenité buňky (švestková pecka, třešňová pecka) nebo do šťavnaté dužniny (citron).
Všechny tři zóny jsou jasně rozlišitelné. Například v plodu třešně je tenká kožovitá vnější vrstva - exokarp, jedlá šťavnatá dužina plodu je mezokarp, tvrdá pecka kamenité tkáně obklopující jediné semeno - endokarp.
Nezralé plody mají většinou zelenou barvu, buňky exokarpu a mezokarpu obsahují chlorofyl a fotosyntetizují. Dozráváním ovoce ztrácí zelenou barvu a často získává jasnou barvu, způsobenou buď antokyany v buněčné šťávě (třešně, hrozny, borůvky) nebo chromoplasty (jeřabina, rajče).
U některých rostlinných druhů se plod tvoří nejen z vaječníku pestíku, ale také z nádobky nebo periantu. Takové plody se nazývají falešné. Například nepravý plod jahodníku ve tvaru bobulí je přerostlá šťavnatá barevná nádoba, na které jsou malé suché skutečné plody (nažky), vytvořené z vaječníků pestíků. Z přerostlé nádoby se tvoří plody šípků, jabloní a hrušek.
U některých druhů rostlin se plod tvoří z několika pestíků. Takovému ovoci se říká prefabrikované, nebo komplexní (složené ovoce maliny, ostružiny, blatouchy).
4. Klasifikace ovoce. Veškerá rozmanitost ovoce je rozdělena do následujících typů.
Nevýrazné suché ovoce
1. Ořech nebo ořech má dřevnaté oplodí obsahující jedno volné semeno. Například v lísce, dubu, buku, konopí.
2. Nažka má kožovité oplodí, které obsahuje jedno volně poléhající semeno. Například u slunečnice, pampelišky, heřmánku a dalších druhů z čeledi hvězdnicovitých.
3. Obilka má kožovité oplodí, které obsahuje jedno semeno srostlé s oplodím. Například u pšenice, ječmene a dalších druhů z čeledi obilnin.
4. Perutýn má oplodí ve tvaru ořechu nebo nažky, které se rozrostlo do jednoho nebo několika okřídlených přívěsků. Například javor, bříza, jilm, jasan.
5. U některých druhů má suchý, neopadavý plod mouchu - chomáč chlupů. Například pampeliška.
Otevírání suchého ovoce
1. Leták - krabicovitý, jednolokulární, vícesemenný plod, tvořený jedním plodolistou, ústící podél ventrálního švu od vrcholu k základně.
Například u bavlníků, druhů z čeledi pryskyřníkovitých a čeledi růžovitých. Pivoňka vytváří mnoho lístků z pestíků jednoho květu.
2. Bob - krabicovitý, jednolokulární, často vícesemenný plod, tvořený jedním plodolistou, otevírající se podél ventrálních a dorzálních švů od vrcholu k základně. Například u hrachu a dalších druhů z čeledi luštěninových. U některých druhů se při otevření fazolových chlopní spirálovitě stáčejí a rozhazují semena (například u žluté akácie). U některých druhů jsou fazole jednosemenné (například u vičence) a pak se neotevírají.
3. Lusk a silique jsou krabicovitý, dvoumístný, vícesemenný plod tvořený dvěma plodolisty. Uprostřed plodu odshora dolů je blanitá přepážka, na jejíž okraje jsou připevněna semena. Ovoce se otevírá dvěma chlopněmi podél dvou švů od základny k vrcholu. Ventily odpadnou, ale přepážka se semínky zůstane. Lusk je dlouhý a úzký plod - délka je 4krát nebo vícekrát větší než šířka (například hořčice) a lusk je krátký a široký (například pastýřská měšec).
4. Tobolka - vícelokulární (zřídka unilokulární) vícesemenný plod tvořený několika plodolisty. Krabice se otevírají různě, buď chlopněmi (například u bavlny, durmanu, vláknitého lnu, čaje, lilií, ricinových bobů), nebo se zoubky na horní straně krabice (například u mnoha druhů z čeledi hřebíčkovitých), nebo s otvory (například u některých druhů máku a zvonu), buď s víkem na horní straně krabice (například v kurníku), nebo s bočními štěrbinami (například v okra).
U některých druhů se suché vícesemenné plody (lusky, fazole) ve zralosti příčně rozpadají na jednosemenné segmenty (například u plané ředkve). Takové plody se nazývají artikulované.
U jiných druhů se suché vícesemenné plody při dozrávání podélně rozpadají na jednotlivé jednosemenné segmenty, např. u javoru a dalších druhů z čeledi javorovitých, dále u čeledí Apiaceae, Lamiaceae, Malvaceae a některých dalších. Takové plody se nazývají zlomkové.
Šťavnaté ovoce
1. Bobule je šťavnatý, obvykle barevný, vícesemenný plod tvořený jedním nebo více plodolisty. Semena v bobulích jsou uložena ve šťavnaté dužině. Například hrozny, rybíz, angrešt, belladonna, borůvky, brusinky, brambory, rajče, okurka, meloun, meloun, dýně, citron, mandarinka, pomeranč (okurka, dýně, meloun, meloun jsou falešné bobule, protože vnější část dužnaté plody, které vytvořily z nádoby).
2. peckovice - obvykle šťavnatý a barevný, jednosemenný, jednosemenný nebo vícesemenný plod, tvořený jedním nebo více plodolisty. Endokarp peckovice má vzhled kamene. Například švestky, třešně, meruňky, dříny, olivy. Vícekamenné peckovice - v černém bezu, řešetláku projímavém. Plodem vlašského ořechu je peckovice, ale jeho mezokarp není šťavnatý; samotný tzv. ořech je semenem plodu. Mezokarp mandlové peckovice také není šťavnatý, zatímco peckovice kokosové palmy je vláknitý.
Plodnost krytosemenných rostlin je velmi vysoká. Jedna rostlina bílé quinoa tedy produkuje asi 100 000 semen ročně, kurník - ≈ 500 000, quinoa - ≈ 750 000,
topoly ≈ 27 000 000 semen.
5. Neplodnost. Distribuce ovoce a semen.
U některých druhů se plod tvoří z květenství. Tento útvar se nazývá neplodnost. Například plodem řepy je kulička, plodem moruše je „bobule“, ovocem vinné révy je „bobule“. Neplodnost moruše jsou srostlé nepravé plody tvořené přerostlými okvětními lístky.
U některých odrůd kulturních rostlin se plody vyvíjejí bez opylení a oplodnění, a tedy bez semen. Tento jev tvorby plodů bez semen se nazývá partenokarpie. Partenokarpie je pozorována u některých odrůd jabloně, hrušně („bez pecek“), hroznů (rozinky – sušené hrozny bez pecek), angreštu, mandarinek, pomerančů, citronů, fíků, japonského tomelu atd. U některých odrůd může být partenokarpie způsobena např. podráždění blizny cizím pylem (opylení hrušky pylem jablek, rajčete pylem brambor, lilku pylem rajčat), některými chemikáliemi, bodnutím hmyzem, pálením horkým drátem apod. Plody bez pecek bývají menší než plody se semeny a proto produkují menší výnosy. V praxi jsou však plody bez pecek spotřebiteli velmi ceněné. Bezsemenné odrůdy se mohou množit pouze vegetativně.
Distribuce ovoce a semen
Semena a plody krytosemenných rostlin vykazují mnoho různých adaptací pro šíření. Většina rostlinných druhů se přizpůsobila šíření semen a plodů větrem (anemochory). Semena bavlny, osiky, topolu, vrby, plodů pampelišky atd. mají zvláštní dlouhé chloupky a vločky, které usnadňují jejich šíření větrem, často na velké vzdálenosti. Plody javoru, břízy, jilmu, jasanu, olše, semena borovice, smrku aj. mají křídlaté výrůstky (proto se plodům říká perutýn), které usnadňují jejich šíření větrem. K šíření větrem přispívá i krycí list květenství lípy. Semena mnoha druhů z čeledí vřesovců, hořců, orchidejí atd. jsou tak malá a lehká, že je vítr unáší jako prach na velmi velké vzdálenosti. Rostliny velbloudí trávy, kurai, ploštice apod., které mají kulovitý tvar, jsou po dozrání plodů odlamovány větrem z podzemní části a valí se po stepi na velké vzdálenosti, často tvoří celé pohyblivé šachty, rozptylují se jejich semena po cestě. Tyto rostliny jsou běžně známé jako tumbleweeds.
Semena a plody vodních rostlin jsou rozptýleny vodou (hydrochory). Dešťová voda (zejména po dešti), voda z potoků a řek se významně podílí na přenosu semen a plodů rostlin pěstovaných na souši.
Zvířata hrají hlavní roli v šíření semen. Šťavnaté plody požírají ptáci (ornithochory) a v menší míře i další živočichové (zoochory). Jejich semena, procházející trávicím traktem zvířete, nejsou poškozena a neztrácejí svou životaschopnost. Spolu se zvířecími exkrementy semena padají na zem a klíčí.
Semena tak často klíčí ve značné vzdálenosti od mateřské rostliny. U některých druhů rostlin (lopuch, mrkev, lopuch, suchý zip) mají semena a plody různé uchycení nebo přilnavé plochy a ulpívají na srsti procházejícího zvířete nebo se lepí na peří ptáků a jsou tak transportovány. Malá semínka jsou roznášena mravenci a mohou se také přilepit na kopyta zvířat v hlíně a být takto nesena.
Některé druhy rostlin si vyvinuly schopnost samostatně rozptýlit svá semena (autochorie). Ve žluté akácii se tedy ventily zralých fazolí otevírají a spirálovitě se točí takovou silou a rychlostí, že se semena přítomná ve fazoli rozptýlí silou do všech směrů. Toto otevírání plodu u žlutého akátu se vysvětluje tím, že při dozrávání jeho bobu se vnější a vnitřní vrstva jeho chlopní nerovnoměrně stahují a mezi nimi vzniká a roste napětí, které způsobuje kroucení a chvění chlopní při otevírání bobu.
V „šílené“ okurce, jejíž zralé plody se odlomí od stopky a semena jsou vyhozena do výsledné díry spolu s tekutým obsahem. To se děje s takovou silou, že semena obvykle padají několik metrů daleko. Toto vymrštění tekutiny a semen se vysvětluje tím, že jak ovoce dozrává, tlak se zvyšuje a v době, kdy ovoce dozrává, dosahuje vysokého napětí.
TÉMA LEKCE 27 : Pryskyřníkovité (Ranunculaceae). Rodina máků (Papaveracea)
1. Čeleď Ranunculaceae. 2. Maková rodina. S. 379 – 383
Řád Ranunculaceae.čeleď Ranunculaceae (2.000)
Asi 2000 druhů. Druhy čeledi pryskyřníkovitých jsou běžné v chladných mírných a subtropických (horských) oblastech. V oblasti Smolensk roste 32 druhů. V tropech se rostliny této čeledi vyskytují výjimečně. Formy života jsou primárně vytrvalé bylinné rostliny, keře, trpasličí keře a vinná réva jsou vzácné.
Listy jsou jednoduché, bez palistů; Tvar listové desky je různý. Listy mohou být buď celé (jarní tráva), nebo v různé míře rozřezané (sasanka lesní, játrovek ušlechtilý, kolumbína). Uspořádání listů je pravidelné.
Mnoho pryskyřníků má upravené podzemní výhony (oddenek sasanky a koňské šišky jarní jasné).
Květy mohou být jednotlivé (rod sasanka) nebo shromážděné v květenstvích (rod pryskyřník, kolumbina, chrpa). Pryskyřníkovité jsou extrémně rozmanité ve struktuře květů.
Rozmanitost ve struktuře květů lze vyjádřit sledováním několika evolučních sérií.
1. Květy s neurčitým počtem členů (u sasanky může mít okvětník 5-8 listů) → květy s určitým, jasně stanoveným počtem členů (rod bojovnice).
2. Okvětník jednoduchý (rod sasanka, rod columbine) → dvoukřídlý (rod Ranunculus)
3. plodolisty z velkého počtu na jeden
4. gynoecium z apokarpního do synkarpního
5. květy od akinomorfních květů po zygomorfní (rod wrestler, rod akonite
6. osa květu je protáhlá, takže květní členy jsou uspořádány do spirály → květní osa je téměř plochá, takže květní členy jsou uspořádány do kruhu.
7. květy jsou nespecializované, entomofilní (často opylované mouchami) → květy jsou specializované, entomofilní (rod wrestler, rod Larkspur).
Mnoho druhů pryskyřníků se vyznačuje tím staminody- vysoce redukované a upravené okvětní lístky (nebo tyčinky), které se mění v nektary. Pouze rostlina bazalka nemá žádné nektary.
Většina druhů má pestíků mnoho, jsou volné a umístěné na protáhlé ose (rody pryskyřník, jaterník, měsíček). Méně často je počet pestíků omezen: 3-1. Vaječník je vždy unilokulární a nadřazený. Obvykle je mnoho vajíček, ale existují druhy, které mají v pestíku 1 vajíčko. Plody jsou letáky nebo ořechy.
Tyčinek může být různý počet. Nejčastěji se jedná o květy s velkým a neurčitým počtem tyčinek, u některých druhů může být od 3 do 1 tyčinek.
měsíček bahenní – * Р 5 А ¥ G ¥
V závislosti na druhu ovoce se v čeledi Ranunculaceae rozlišují dvě podčeledi:
· zimníky (rody plavky, zápasník, skřivan), mající letákové ovoce
· Pryskyřníkovité (rody Pryskyřník, Anemone, Bazalka), mající ořechové plody
Alkaloidy jsou velmi běžné ve všech rostlinách této čeledi. Pryskyřníky jsou jedovaté, nejsou to živné rostliny, ale pro množství alkaloidů se rostliny používají jako léčiva. U morfologicky nespecializovaných druhů (rod bazalka, měsíček a pryskyřník) se syntetizují jednoduché alkaloidy, u pokročilých a specializovaných rodů komplexní alkaloidy (rostliny z rodu akonit a skřivan).
Z různých druhů pryskyřníků se v kardiologické praxi používá 20 alkaloidů. Alkaloidy všech druhů pryskyřníků jsou jedovaté, ty nejjedovatější patří do čeledi akonitů.
Jarní hlaváček, skřivan síťkovaný a borax (neboli akonit) jsou důležité léčivé rostliny.
Alkaloidy v jarním adonisu vyvolávají účinek podobný kurare a používají se k uvolnění svalů.
Alkaloidy larkspur reticularis se používají v nervové klinice při onemocněních spojených s poruchami motorických funkcí
Pro radikulitidu se doporučuje tinktura z byliny džungarského akonitu, která je součástí léku "Akofit".
Vzácné a chráněné druhy: plavec evropský, vrána ostnitá, zápasník severní.
Objednejte Poppy.Poppy family (250)
Rostliny této čeledi jsou rozšířeny především v tropických a subtropických oblastech. Vzácně se vyskytuje v mírném pásmu. Na území regionu Smolensk se vyskytují 2 druhy - vlaštovičník větší a mák.
Mák - bylinné vytrvalé, někdy jednoleté rostliny. Přítomnost letniček je považována za evolučně mladou vlastnost. Uspořádání listů je střídavé, listy jsou jednoduché, bez palistů, mohou být buď celé, nebo rozřezané.
Květy máku jsou často velké a aktinomorfní:
* Ca 2 Co 2 +2 A ¥ G ( ¥ )
Sepaly zpravidla opadávají, když se květ otevře. Gynoecium se skládá z několika plodolistů, které srůstají a vytvářejí parakarpní gynoecium. Plodem je suchá tobolka obalená hřebíčkem. Existuje mnoho vajíček, plody jsou malé a obsahují tuky jako rezervní materiál.
Nápadným anatomickým a biochemickým znakem máku je přítomnost laticiferů, ve kterých se syntetizuje mléčná míza složité struktury. Obsahuje různé isochinolinové alkaloidy. Mléčná míza a alkaloidy máku se široce používají v lékařství.
Zvláštní význam nabyl rod mák, uspávající druh máku je hlavní surovinou pro výrobu opia. Opium obsahuje více než 20 alkaloidů, včetně morfinu, kodeinu, narkotinu a papaverinu. Některé alkaloidy mají silný anestetický účinek, ale jsou návykové. Papaverin se používá jako spazmolytikum při angíně pectoris a bronchiálním astmatu. U nás se mák uspávající nepěstuje. Určité množství omamných alkaloidů obsahují také olejniny a okrasné odrůdy máku.
Skvělý celandine
Květy jsou malé, shromážděné v deštníkovitém květenství: * Ca 2 Co 2 +2 A ¥ G (¥)
Jeho pomerančová mléčná šťáva spolu s alkaloidy obsahuje flavonoidy, třísloviny, saponiny, organické kyseliny a vitamíny. Vlaštovičník se používá jako choleretikum a antibakteriální prostředek při onemocněních jater a žlučníku. Vlaštovičník větší je jedovatá rostlina.
Droga "Sangviritrin" se vyrábí z byliny vlaštovičník pro vyplachování úst a krku při zánětu a také pro mytí hnisavých ran.
Žlutý mák se používá k léčebným účelům a používá se k získávání Ne narkotické antitusikum glaucin hydrochlorid. K léčebným účelům se využívá i makaka maloplodá a makaka srdčitá.
LEKCE 28: čeleď Rosaceae.
1. Čeleď Rosaceae, obecná charakteristika.
2. Podčeleď Spiraea. Podčeleď růžová.
3. Podrodina Apple. Podčeleď švestek.
1. čeleď Rosaceae (3500)
Zástupci této rodiny jsou distribuováni mimo tropické oblasti severní polokoule a nacházejí se v různých fytocenózách. V oblasti Smolensk roste 56 druhů Rosaceae. Životní forma: stromy, keře, byliny (jednoleté i víceleté).
Listy jsou jak jednoduché (celé nebo členité), tak i složité (párové i nepárnopeřené, trojčetné), velmi často se vyskytují palisty. Uspořádání listů je střídavé, zřídka opačné. Některé druhy mají trny (metamorfózy epidermis).
Biochemické vlastnosti Rosaceae:
· nedochází k nespecializovanému sekundárnímu metabolismu. To se projevuje úplnou nepřítomností alkaloidů a toxinů. Esenciální oleje vznikají zřídka a vždy mají jednoduché složení a strukturu (s výjimkou rodu Rose)
Struktura květů Rosaceae není příliš specializovaná na opylování hmyzem. Květy jsou jednotlivé nebo shromážděné v květenstvích různých typů. Květy jsou aktinomorfní, okvětní lístek je složitý - bývá pět kališních lístků a okvětních lístků. Dost často má květina dvojnásobný počet kališních lístků, v takovém případě se tvoří první kruh kališních lístků nižší. Existuje mnoho tyčinek, 2-4krát více než okvětních lístků. Počet plodolistů je buď neurčitý, nebo přísně fixní.
Charakteristickým znakem květiny je přítomnost hypanthia- zvláštní útvar tvořený zarostlou schránkou a s ní srostlými základy kališních lístků, okvětních lístků a tyčinek. Tvar hypanthia může být konvexní, talířovitý nebo konkávní. Hypanthium chrání gynoecium před vnějšími faktory. Zároveň hraje důležitou roli při tvorbě plodů. Může růst a podílet se na tvorbě mezokarpů a endokarpů. Šťavnatá dužina šípků, jabloní, hrušek a švestek je tedy přerostlé hypanthium. Kromě toho hypanthium zajišťuje komunikaci se zvířaty, která roznášejí plody (exo- a endochorie).
Plody Rosaceae jsou velmi rozmanité ve struktuře, což poskytuje různé způsoby distribuce. Zdokonalování metod distribuce je jedním z hlavních směrů evoluce v rámci rodiny.
Rosaceae velmi často produkují falešné plody (jablko v jabloni a jablko v horském jasanu) a komplexní ovoce (více ořechů v jahodách a mnohodrupe v malinách).
Čeleď Rosaceae zahrnuje čtyři podčeledi: spireaceae, roseaceae, jablko a švestku.
2. Podčeleď Spiraea. Podčeleď růžová.
Podčeleď Spiraea (180)
Jedná se o nejprimitivnější podčeleď, kterou představují keře, méně často stromy a velmi vzácně vytrvalé oddenkové byliny. Květy jsou drobné, sbírané buď v latách, nebo v hroznovitých květenstvích nebo v chocholících. Okvětní lístky koruny jsou bílé, méně často světle růžové. Zástupci této čeledi se vyznačují konkávní nebo téměř plochou nádobou (hypanthium) a plody jsou suché - vícelisté (kvintalisté). Gynoecium je apokarpní, obvykle sestává z 2-5 plodolistů.
Čeleď zahrnuje rody Spirea, Meadowsweet a Jeřabina.
Ústředním rodem je rod Spiraea, rozšířený na Sibiři a Dálném východě. Spiraea jsou keře s jednoduchými listy, bez palistů. Květy jsou obvykle bílé nebo růžové, v latovitých, korymbózních nebo deštníkovitých květenstvích. Plod je vícelistý.
Rod luční, druh luční. Jedná se o velkou vytrvalou rostlinu vysokou až 2 m s krátkým oddenkem. Listy jsou přerušovaně lichozpeřené, svrchu tmavě zelené, dole s tenkým bíle plstnatým dospíváním. Při tření listy vydávají štiplavý zápach. Květy jsou žlutavě bílé a shromážděné v hustých panikulovitých květenstvích. Plody jsou jednosemenné lístečky. V lidovém léčitelství se odpradávna používala lučina jako adstringens.
* Ca (5) Co 5 A ¥ G 6-10
Podčeleď Rosaceae (1 700)
Bylinné rostliny jsou rozšířeny v různých fytocenózách od tundry až po horské tropy. Květy se sbírají v corymbose nebo racemose dichasia, méně často osamělých květinách.
psí růže – * Ca (5) Co 5 A ¥ G ¥
Gynoecium je apokarpní, polymerní, ale vaječník obsahuje vždy jedno, zřídka dvě vajíčka.
Plody jsou rozmanité, ale nejčastěji se jedná o agregované peckovice a víceoříšky. Mnoho druhů této čeledi má šťavnaté komplexní plody, na jejichž tvorbě se podílí hypanthium.
Mnoho rostlin této čeledi se vyznačuje rozmnožováním pomocí oddenků, stolonů nebo úponků.
Jako léčiva se používají látky sekundárního metabolismu růží.
1. maliník obecný - jeho plody obsahují značné množství kyseliny salicylové, která podmiňuje jejich diaforetický a antipyretický účinek proti nachlazení.
2. palina - květ nemá korunu, kalich je fialový a 4dílný. Odvar z oddenku se používá jako hemostatikum při děložním a hemoroidním krvácení.
3. Potentilla erecta (Uzik, Kalgan). Vyznačuje se 4členným okvětím a kalichem s podkalichem. Galangalová tinktura (25g oddenku na 500ml vodky) se používá při střevních katarech a úplavici.
4. mochna bahenní (lidový název - dekop) - odvar z oddenku se používá při kloubních onemocněních a žloutence. Používá se také jako adstringens, hemostatikum a diaforetikum.
5. Skořice šípková – obsahuje velké množství vitamínu C, dále vitamíny B2, K, karoten a kyselinu citronovou. Kondenzovaný vodný extrakt ze šípku s cukrem (holosas) se používá jako choleretikum.
3. Podrodina Apple. Podčeleď švestek.
 ODDĚLENÍ ČERVENÝCH ŘAS je téměř výhradně z mořských řas. životní cyklus - úplná absence bičíkových stádií, zvláštní forma oogamního sexuálního procesu. 2
ODDĚLENÍ ČERVENÝCH ŘAS je téměř výhradně z mořských řas. životní cyklus - úplná absence bičíkových stádií, zvláštní forma oogamního sexuálního procesu. 2
 Pigmenty chlorofyl „a“ a „b“, karotenoidy (karoten, zeaxanthin, antheraxanthin, kryptoxantin, lutein, neoxanthin) fykobiliny: fykoerytriny (červené) fykocyaniny a alofykocyanin (modrá) barva ocelově modrého thallusu od ocelově modré po modrou nadbytek fykocyaninu) 3
Pigmenty chlorofyl „a“ a „b“, karotenoidy (karoten, zeaxanthin, antheraxanthin, kryptoxantin, lutein, neoxanthin) fykobiliny: fykoerytriny (červené) fykocyaniny a alofykocyanin (modrá) barva ocelově modrého thallusu od ocelově modré po modrou nadbytek fykocyaninu) 3
 Chloroplasty jsou obalem dvou membrán, jednotlivých tylakoidů, na povrchu fykobilisomu. Rezervním produktem je polysacharid „purpurový škrob“, který získává od jódu hnědočervenou barvu. Fialová škrobová zrna jsou uložena v cytoplazmě vždy bez spojení s pyrenoidy a chromatofory. 4
Chloroplasty jsou obalem dvou membrán, jednotlivých tylakoidů, na povrchu fykobilisomu. Rezervním produktem je polysacharid „purpurový škrob“, který získává od jódu hnědočervenou barvu. Fialová škrobová zrna jsou uložena v cytoplazmě vždy bez spojení s pyrenoidy a chromatofory. 4
 Struktura stélky je jednobuněčné kokoidní formy (porphyridium), heterotrichní a ve formě rozvětvených filament připojených k substrátu pomocí rhizoidů. pseudoparenchymatózní stélky, propletení postranních větví Lamelární stélky parenchymatické struktury (porfyr). 5
Struktura stélky je jednobuněčné kokoidní formy (porphyridium), heterotrichní a ve formě rozvětvených filament připojených k substrátu pomocí rhizoidů. pseudoparenchymatózní stélky, propletení postranních větví Lamelární stélky parenchymatické struktury (porfyr). 5
 Buňka je pokryta pektinovým obalem - hemicelulózové složky bobtnají a ve stěnách se ukládá vápno. Buňky jsou jedno- a vícejaderné.Chromatofory jsou parietální, četné, ve formě zrn nebo destiček. 6
Buňka je pokryta pektinovým obalem - hemicelulózové složky bobtnají a ve stěnách se ukládá vápno. Buňky jsou jedno- a vícejaderné.Chromatofory jsou parietální, četné, ve formě zrn nebo destiček. 6
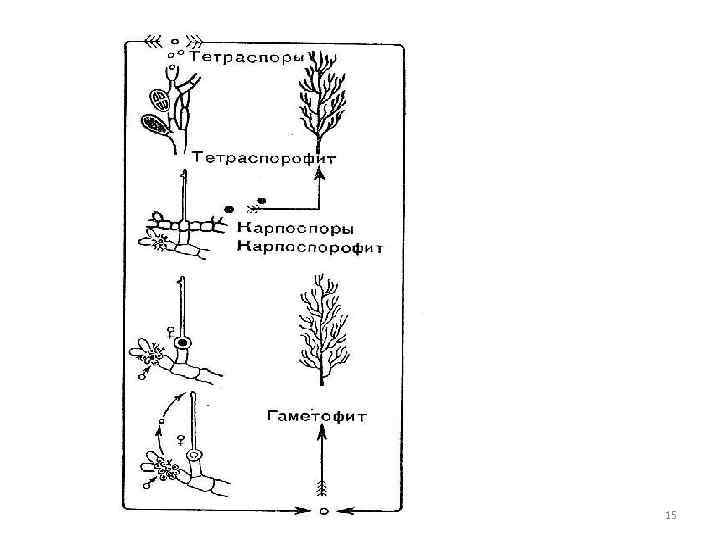
 Nepohlavní rozmnožování přes nepohyblivé buňky monospory - jedna ve sporangiu Tetraspory - čtyři tetraspory - na diploidních nepohlavních rostlinách - sporofytech. U sporangií dochází k meióze před tvorbou tetraspor. 7
Nepohlavní rozmnožování přes nepohyblivé buňky monospory - jedna ve sporangiu Tetraspory - čtyři tetraspory - na diploidních nepohlavních rostlinách - sporofytech. U sporangií dochází k meióze před tvorbou tetraspor. 7
 Pohlavní výběžek Oogamní Ženský orgán - většinou karpogon - z rozšířené bazální části - břicho - vajíčko a výběžek - trichogyna. Carpogon se vyvíjí na speciální krátké karpogoniální větvi. 8
Pohlavní výběžek Oogamní Ženský orgán - většinou karpogon - z rozšířené bazální části - břicho - vajíčko a výběžek - trichogyna. Carpogon se vyvíjí na speciální krátké karpogoniální větvi. 8
 Antheridia - malé bezbarvé buňky obsahující bičíkovité spermie, spermie jsou pasivně transportovány vodními proudy a přilnou k trichogynu. V místě kontaktu se spermie a trichogyny rozpouštějí jejich stěny, jádro spermie se pohybuje po trichogynu v břišní části karpogonu, splývá s 9
Antheridia - malé bezbarvé buňky obsahující bičíkovité spermie, spermie jsou pasivně transportovány vodními proudy a přilnou k trichogynu. V místě kontaktu se spermie a trichogyny rozpouštějí jejich stěny, jádro spermie se pohybuje po trichogynu v břišní části karpogonu, splývá s 9
 tvorba karpospor Po oplození je bazální část karpogonu oddělena přepážkou od trichogyna - odumírá Vývoj - tvorba karpospor obsah zygoty (oplodněného karpogonu) 1. přímo se dělí za vzniku nepohyblivých holých spor - karpospor, 10
tvorba karpospor Po oplození je bazální část karpogonu oddělena přepážkou od trichogyna - odumírá Vývoj - tvorba karpospor obsah zygoty (oplodněného karpogonu) 1. přímo se dělí za vzniku nepohyblivých holých spor - karpospor, 10
 tvorba karpospor 2. z oplodněného karpogonu vyrůstají větvící vlákna - gonimoblasty, jejich buňky se mění v karposporangia, produkují vždy jednu karposporu. jedenáct
tvorba karpospor 2. z oplodněného karpogonu vyrůstají větvící vlákna - gonimoblasty, jejich buňky se mění v karposporangia, produkují vždy jednu karposporu. jedenáct
 tvorba karpospor ve většině gonimoblastů se nevyvíjí přímo z břicha oplodněného karpogonu z acoxylárních buněk. mohou být odstraněny z karpogonu nebo jsou umístěny na stélce v těsné blízkosti, když jsou z karpogonu odstraněny pomocné buňky, pojivové buňky nebo oblasti rostou z jeho břicha po oplodnění y e, závity. 12
tvorba karpospor ve většině gonimoblastů se nevyvíjí přímo z břicha oplodněného karpogonu z acoxylárních buněk. mohou být odstraněny z karpogonu nebo jsou umístěny na stélce v těsné blízkosti, když jsou z karpogonu odstraněny pomocné buňky, pojivové buňky nebo oblasti rostou z jeho břicha po oplodnění y e, závity. 12
 buňky oblastícových filament obsahují diploidní jádra. Oblastková filamenta rostou směrem k pomocným buňkám, v místě kontaktu se membrány rozpouštějí a vzniká komunikace mezi buňkou oblasticového vlákna a pomocnou buňkou. Toto splynutí buněk není doprovázeno splynutím jejich jader (diploidního buněčného jádra ooblastického vlákna a haploidního jádra pomocného vlákna). fúze s pomocnou buňkou stimuluje dělení diploidního jádra oblastem filamentové buňky a vývoj gonimoblastů obsahuje diploidní jádra a produkují diploidní karpospory Gonimoblasty - speciální generace - 13
buňky oblastícových filament obsahují diploidní jádra. Oblastková filamenta rostou směrem k pomocným buňkám, v místě kontaktu se membrány rozpouštějí a vzniká komunikace mezi buňkou oblasticového vlákna a pomocnou buňkou. Toto splynutí buněk není doprovázeno splynutím jejich jader (diploidního buněčného jádra ooblastického vlákna a haploidního jádra pomocného vlákna). fúze s pomocnou buňkou stimuluje dělení diploidního jádra oblastem filamentové buňky a vývoj gonimoblastů obsahuje diploidní jádra a produkují diploidní karpospory Gonimoblasty - speciální generace - 13
 U nejorganizovanějších červených řas se pomocné buňky vyvíjejí až po oplození karpogona a v jeho bezprostřední blízkosti. Kombinace pomocné buňky (nebo buněk) s karpogonem se speciálně nazývá prokarp. Odpadá vznik dlouhých ooblastických filament spojujících karpogon a pomocné buňky, pomocná buňka jednoduše splyne s břichem oplodněného karpogonu, načež se z ní vyvinou gonimoblasty s karposporami.Karposporangia se často nacházejí v úzkých skupinách - cystokarpy, pokrytá pseudoparenchymatózní membránou vyvíjející se z buněk sousedících s karpogonem. 14
U nejorganizovanějších červených řas se pomocné buňky vyvíjejí až po oplození karpogona a v jeho bezprostřední blízkosti. Kombinace pomocné buňky (nebo buněk) s karpogonem se speciálně nazývá prokarp. Odpadá vznik dlouhých ooblastických filament spojujících karpogon a pomocné buňky, pomocná buňka jednoduše splyne s břichem oplodněného karpogonu, načež se z ní vyvinou gonimoblasty s karposporami.Karposporangia se často nacházejí v úzkých skupinách - cystokarpy, pokrytá pseudoparenchymatózní membránou vyvíjející se z buněk sousedících s karpogonem. 14
 15
15

 CLASS BANGUY Buňky často s hvězdicovým chromatoforem a pyrenoidem. Póry mezi buňkami obvykle chybí. Carpogon bez trichogynu, po oplodnění se obsah karpogonu přímo rozdělí na karpospory. Nepohlavní rozmnožování - monospory 17
CLASS BANGUY Buňky často s hvězdicovým chromatoforem a pyrenoidem. Póry mezi buňkami obvykle chybí. Carpogon bez trichogynu, po oplodnění se obsah karpogonu přímo rozdělí na karpospory. Nepohlavní rozmnožování - monospory 17

 19
19
 rod compsopogon Sladkovodní rozšířený v tropech zavlečený s akvarijními rostlinami Thallus heterotrichous Rozmnožování monosporami. Monosporangia jsou oddělena šikmou přepážkou od kterékoli buňky stélky a mají zrnitý obsah z nadbytku purpurového škrobu, který se mění v jednu nehybnou nahou monosporu 20
rod compsopogon Sladkovodní rozšířený v tropech zavlečený s akvarijními rostlinami Thallus heterotrichous Rozmnožování monosporami. Monosporangia jsou oddělena šikmou přepážkou od kterékoli buňky stélky a mají zrnitý obsah z nadbytku purpurového škrobu, který se mění v jednu nehybnou nahou monosporu 20
 21
21

 TŘÍDA FLORIDA Buňky většinou s parietálními chromatofory bez pyrenoidů, mezi buňkami jsou póry. Carpogon s trichogynem. Po oplození se gonimoblasty vyvíjejí buď přímo z břicha oplodněného karpogonu, nebo z pomocných buněk po jejich splynutí s oblastcovými vlákny. Nepohlavní rozmnožování většinou probíhá pomocí tetraspor. Gonimoblasty nesoucí karposporangia jsou považovány za zvláštní generaci - karposporofyt. Podrobnosti vývoje karposporofytu (z oplodněného karpogonu nebo z pomocných buněk), stejně jako doba diferenciace pomocných buněk (před nebo po oplození), jejich poloha na stélce atd. tvoří základ pro rozdělení floridia do šesti řádů 23
TŘÍDA FLORIDA Buňky většinou s parietálními chromatofory bez pyrenoidů, mezi buňkami jsou póry. Carpogon s trichogynem. Po oplození se gonimoblasty vyvíjejí buď přímo z břicha oplodněného karpogonu, nebo z pomocných buněk po jejich splynutí s oblastcovými vlákny. Nepohlavní rozmnožování většinou probíhá pomocí tetraspor. Gonimoblasty nesoucí karposporangia jsou považovány za zvláštní generaci - karposporofyt. Podrobnosti vývoje karposporofytu (z oplodněného karpogonu nebo z pomocných buněk), stejně jako doba diferenciace pomocných buněk (před nebo po oplození), jejich poloha na stélce atd. tvoří základ pro rozdělení floridia do šesti řádů 23
 Řád Nemalionaceae Zástupci se vyznačují absencí pomocných buněk, gonimoblasty se vyvíjejí přímo z oplodněného karpogonu 24
Řád Nemalionaceae Zástupci se vyznačují absencí pomocných buněk, gonimoblasty se vyvíjejí přímo z oplodněného karpogonu 24



 Rod Lemanea se vyskytuje v rychle tekoucích řekách se studenou vodou. Thallus má vzhled nevětvících štětin o délce 10-15 cm a tloušťce 1 mm, tmavě fialové nebo olivově hnědé barvy s nodulárními otoky. Přichyceno k podkladu pomocí podešve vyrobené z plazivých nití. Jedna nit protáhlých bezbarvých buněk probíhá podél osy talu. Z vrcholu každé buňky centrální osy se rozprostírá přeslen většinou čtyř radiálních větví. Bazální buňky každé takové větve jsou velké a protáhlé. Z jejich distálního (morfologicky horního) konce vybíhají větve druhého řádu, které se zase větví opakovaně. Koncové větve srůstají do vícevrstvé kůry. Jeho vnější buňky jsou malé a naplněné chromatofory, 28
Rod Lemanea se vyskytuje v rychle tekoucích řekách se studenou vodou. Thallus má vzhled nevětvících štětin o délce 10-15 cm a tloušťce 1 mm, tmavě fialové nebo olivově hnědé barvy s nodulárními otoky. Přichyceno k podkladu pomocí podešve vyrobené z plazivých nití. Jedna nit protáhlých bezbarvých buněk probíhá podél osy talu. Z vrcholu každé buňky centrální osy se rozprostírá přeslen většinou čtyř radiálních větví. Bazální buňky každé takové větve jsou velké a protáhlé. Z jejich distálního (morfologicky horního) konce vybíhají větve druhého řádu, které se zase větví opakovaně. Koncové větve srůstají do vícevrstvé kůry. Jeho vnější buňky jsou malé a naplněné chromatofory, 28
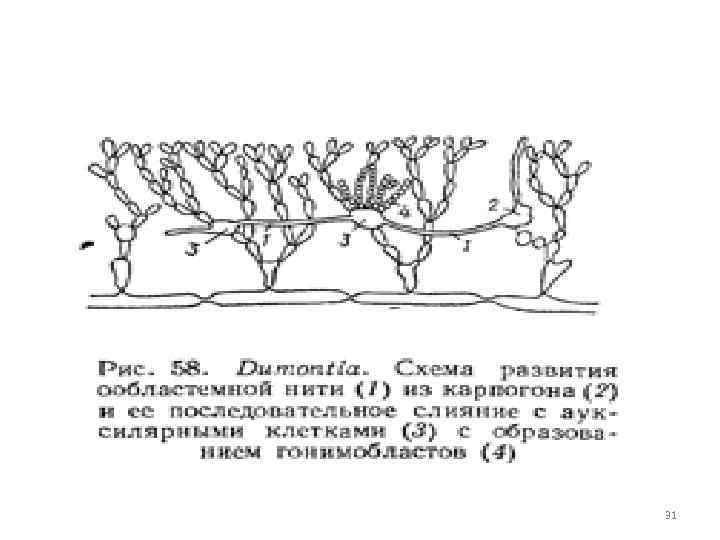
 Řád Cryptonemia obsahuje pomocné buňky, které se vyvíjejí před oplodněním karpogonu a jsou rozptýleny po celém stélku v určité vzdálenosti od karpogonu. Z karpogonu po oplodnění vyrůstají do pomocných buněk více či méně dlouhá mnohobuněčná pojivová neboli ooblastická vlákna. Po splynutí pomocné buňky s buňkou ooblastického filamenta a přechodu diploidního jádra filamentární buňky v pomocnou buňku vznikají gonimoblasty, vyvíjející se na gonimoblastech (karposporofytech), karpospory obsahují diploidní jádro a vyklíčí v diploidní rostliny - tetrasporofyty, které produkují pouze orgány nepohlavního rozmnožování - tetrasporangia. Když se tvoří tetraspory, dochází k meióze a haploidní tetraspory rostou v haploidní gametofytové rostliny nesoucí reprodukční orgány. Gametofyt a tetrasporofyt se zevně (morfologicky) neliší. izomorfní změna generací, komplikovaná diploidní karposporoftózou. 29
Řád Cryptonemia obsahuje pomocné buňky, které se vyvíjejí před oplodněním karpogonu a jsou rozptýleny po celém stélku v určité vzdálenosti od karpogonu. Z karpogonu po oplodnění vyrůstají do pomocných buněk více či méně dlouhá mnohobuněčná pojivová neboli ooblastická vlákna. Po splynutí pomocné buňky s buňkou ooblastického filamenta a přechodu diploidního jádra filamentární buňky v pomocnou buňku vznikají gonimoblasty, vyvíjející se na gonimoblastech (karposporofytech), karpospory obsahují diploidní jádro a vyklíčí v diploidní rostliny - tetrasporofyty, které produkují pouze orgány nepohlavního rozmnožování - tetrasporangia. Když se tvoří tetraspory, dochází k meióze a haploidní tetraspory rostou v haploidní gametofytové rostliny nesoucí reprodukční orgány. Gametofyt a tetrasporofyt se zevně (morfologicky) neliší. izomorfní změna generací, komplikovaná diploidní karposporoftózou. 29
 Durenea rod běžný v jižních mořích. Talus vypadá jako rozvětvené, slizké, růžové keře. třicet
Durenea rod běžný v jižních mořích. Talus vypadá jako rozvětvené, slizké, růžové keře. třicet
 31
31
 32
32
 33
33


 Řád Ceramiaceae je vysoce organizovaný, mnoho druhů pomocných buněk se diferencuje až po oplození karpogonu Nevytvářejí se dlouhá vlákna oblasti: břicho karpogonu buď přímo splyne se sousední pomocnou buňkou (buňkami), nebo krátkými procesy , tvořící cystokarpy 36
Řád Ceramiaceae je vysoce organizovaný, mnoho druhů pomocných buněk se diferencuje až po oplození karpogonu Nevytvářejí se dlouhá vlákna oblasti: břicho karpogonu buď přímo splyne se sousední pomocnou buňkou (buňkami), nebo krátkými procesy , tvořící cystokarpy 36


 evoluce v rámci oddělení Banguiaceae je primitivnější. Carpogon ještě nemá vyvinutou typickou formu a od běžných vegetativních buněk se liší jen málo. Po oplodnění se obsah karpogonu přímo rozdělí na karpospory. Floridai je vyvinutější skupina Carpogon s Trichogyna. Z třídy floridií je nejjednodušším řádom nemalionaceae, které nemají pomocné buňky a přímo z břišní části oplodněného karpogonu se tvoří gonimoblasty, na kterých se vyvíjejí karposporangia. Kryptonémie jsou dalším krokem v progresivní evoluci: mají pomocné buňky, které přispívají ke zvýšení produkce karpospor, protože nevzniká jeden cystokarp, ale mnoho - podle počtu pomocných buněk. Když jsou pomocné buňky náhodně rozptýleny po stélce, je zapotřebí více či méně dlouhých oblastícových vláken. Nejvyššího stupně evoluce dosáhly ceramiidy, které mají prokarp a pomocné buňky se diferencují až po oplození. Těsná blízkost karpogonu a pomocné buňky (buňek) v prokarpu usnadňuje tvorbu cpstokarpů. Tento řád je druhově nejbohatší. 39
evoluce v rámci oddělení Banguiaceae je primitivnější. Carpogon ještě nemá vyvinutou typickou formu a od běžných vegetativních buněk se liší jen málo. Po oplodnění se obsah karpogonu přímo rozdělí na karpospory. Floridai je vyvinutější skupina Carpogon s Trichogyna. Z třídy floridií je nejjednodušším řádom nemalionaceae, které nemají pomocné buňky a přímo z břišní části oplodněného karpogonu se tvoří gonimoblasty, na kterých se vyvíjejí karposporangia. Kryptonémie jsou dalším krokem v progresivní evoluci: mají pomocné buňky, které přispívají ke zvýšení produkce karpospor, protože nevzniká jeden cystokarp, ale mnoho - podle počtu pomocných buněk. Když jsou pomocné buňky náhodně rozptýleny po stélce, je zapotřebí více či méně dlouhých oblastícových vláken. Nejvyššího stupně evoluce dosáhly ceramiidy, které mají prokarp a pomocné buňky se diferencují až po oplození. Těsná blízkost karpogonu a pomocné buňky (buňek) v prokarpu usnadňuje tvorbu cpstokarpů. Tento řád je druhově nejbohatší. 39
 Nejstarší fosílie identifikovaná jako červená řasa je také nejstarší eukaryotickou fosílií patřící do specifického moderního taxonu. Bangiomorpha pubescens, mnohobuněčná fosilie nalezená v arktické Kanadě, se jen mírně liší od moderních červených řas rodu Bangia, přestože byla uložena ve vrstvách před 1,2 miliardami let. 41
Nejstarší fosílie identifikovaná jako červená řasa je také nejstarší eukaryotickou fosílií patřící do specifického moderního taxonu. Bangiomorpha pubescens, mnohobuněčná fosilie nalezená v arktické Kanadě, se jen mírně liší od moderních červených řas rodu Bangia, přestože byla uložena ve vrstvách před 1,2 miliardami let. 41
 DIVIZE HNĚDÉ ŘASY RNAEORNUTH mořské, zvláště hojné ve studených vodách severní a jižní polokoule. Z hlediska morfologické a anatomické diferenciace thallusu je na vyšší úrovni než dříve uvažované skupiny řas. Nejsou známy ani jednobuněčné ani koloniální formy, ani stélka ve formě jednoduchého nerozvětveného vlákna. nejjednodušší jsou heterotrichní stélky, velké, s falešnou nebo pravou tkáňovou strukturou.
DIVIZE HNĚDÉ ŘASY RNAEORNUTH mořské, zvláště hojné ve studených vodách severní a jižní polokoule. Z hlediska morfologické a anatomické diferenciace thallusu je na vyšší úrovni než dříve uvažované skupiny řas. Nejsou známy ani jednobuněčné ani koloniální formy, ani stélka ve formě jednoduchého nerozvětveného vlákna. nejjednodušší jsou heterotrichní stélky, velké, s falešnou nebo pravou tkáňovou strukturou.
 silně slizovité buněčné stěny, jedno jádro, jedna nebo více vakuol, obvykle stěnové chromatofory různých tvarů. chromatofory jsou obklopeny složitým systémem membrán - v přímé souvislosti s jaderným obalem - „chloroplastové endoplazmatické retikulum“ Matrice chloroplastu je protnuta paralelními tritylakoidními lamelami Pyrenoid bez thylakoidů vyčnívá z chloroplastu ve formě ledviny
silně slizovité buněčné stěny, jedno jádro, jedna nebo více vakuol, obvykle stěnové chromatofory různých tvarů. chromatofory jsou obklopeny složitým systémem membrán - v přímé souvislosti s jaderným obalem - „chloroplastové endoplazmatické retikulum“ Matrice chloroplastu je protnuta paralelními tritylakoidními lamelami Pyrenoid bez thylakoidů vyčnívá z chloroplastu ve formě ledviny
 Chromatoforové pigmenty jsou zbarveny do hněda, chlorofyly „a“ a „c“ (chlorofyl „b“ chybí) β-karoten Existuje mnoho hnědých xantofylů, zejména fukoxanthin. 44
Chromatoforové pigmenty jsou zbarveny do hněda, chlorofyly „a“ a „c“ (chlorofyl „b“ chybí) β-karoten Existuje mnoho hnědých xantofylů, zejména fukoxanthin. 44
 Rezervní polysacharid - laminarinalkohol mannitolové tuky - se ukládá mimo chloroplast v cytoplazmě. Monádové buňky (zoospory a gamety) mají oko a bičíky. Ocellus je součástí plastidu a je spojen s bičíkovým aparátem. Bičíky jsou heteromorfní. 45
Rezervní polysacharid - laminarinalkohol mannitolové tuky - se ukládá mimo chloroplast v cytoplazmě. Monádové buňky (zoospory a gamety) mají oko a bičíky. Ocellus je součástí plastidu a je spojen s bičíkovým aparátem. Bičíky jsou heteromorfní. 45


 Rozmnožování je vegetativní, nepohlavní a pohlavní. Vegetativní množení sekcemi stélky. Některé mají specializované větve (plodové pupeny), které se lámou do nových stélků. Nepohlavní rozmnožování - zoospory vzniklé v jednobuněčných nebo jednokomorových výtrusnicích na diploidních rostlinách (sporofytech) před vznikem zoospor, jádro je redukovaně rozděleno Haploidní zoospory - na haploidní pohlavní rostliny - gametofyty, na kterých se tvoří pohlavní orgány. U prvoků je pohlavní proces izogamní, gamety se vyvíjejí v multilokulárních nebo vícekomorových sporangiách, u nejorganizovanějších hnědých řas je pohlavní proces oogamní. V oogonii a antheridii se zpravidla tvoří jedna gameta (vajíčko a spermie). Vajíčko je vždy oplodněno mimo oogium. Zygota roste v diploidní rostlinu bez období klidu.
Rozmnožování je vegetativní, nepohlavní a pohlavní. Vegetativní množení sekcemi stélky. Některé mají specializované větve (plodové pupeny), které se lámou do nových stélků. Nepohlavní rozmnožování - zoospory vzniklé v jednobuněčných nebo jednokomorových výtrusnicích na diploidních rostlinách (sporofytech) před vznikem zoospor, jádro je redukovaně rozděleno Haploidní zoospory - na haploidní pohlavní rostliny - gametofyty, na kterých se tvoří pohlavní orgány. U prvoků je pohlavní proces izogamní, gamety se vyvíjejí v multilokulárních nebo vícekomorových sporangiách, u nejorganizovanějších hnědých řas je pohlavní proces oogamní. V oogonii a antheridii se zpravidla tvoří jedna gameta (vajíčko a spermie). Vajíčko je vždy oplodněno mimo oogium. Zygota roste v diploidní rostlinu bez období klidu.

 TŘÍDA ISOGENERATES Řád Ectocarpous Rod Ectocarpus - nejprimitivnější hnědá řasa. Jsou rozšířeny ve všech mořích, zejména studených, a rostou na podmořských předmětech a jiných větších řasách. 50
TŘÍDA ISOGENERATES Řád Ectocarpous Rod Ectocarpus - nejprimitivnější hnědá řasa. Jsou rozšířeny ve všech mořích, zejména studených, a rostou na podmořských předmětech a jiných větších řasách. 50
 51
51
 52
52

 54
54
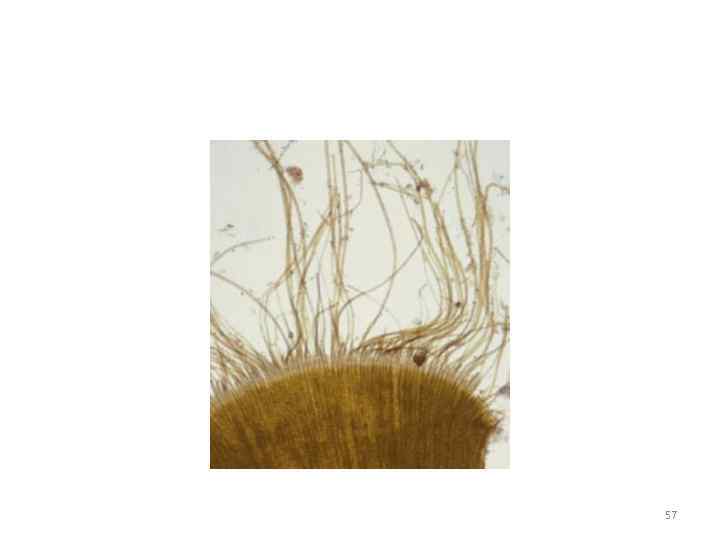
 Řád Cutleriaeae Řád zahrnuje pouze dva rody: Cutleria a Zanardinia Cutleria je rozšířena podél celého pobřeží Evropy, Zanardinie - především ve Středozemním moři a také v Černém moři. Oba rody vykazují střídání generací: u Cutleria je změna generací heteromorfní, v Zanardinii je izomorfní 55
Řád Cutleriaeae Řád zahrnuje pouze dva rody: Cutleria a Zanardinia Cutleria je rozšířena podél celého pobřeží Evropy, Zanardinie - především ve Středozemním moři a také v Černém moři. Oba rody vykazují střídání generací: u Cutleria je změna generací heteromorfní, v Zanardinii je izomorfní 55

 57
57
 58
58

 60
60

 62
62
 TŘÍDA HETEROGENERÁTNÍ Řád Laminaria Gametofyty se od sebe málo liší a jsou zastoupeny mikroskopickými, často na několik buněk redukovanými, vláknitými rostlinami nesoucími rozmnožovací orgány. Na samčích gametofytech se tvoří antheridia ve formě malých buněk, které vyvíjejí jednu spermii, na samičích gametofytech - oogonia, ve kterých se tvoří jedno vajíčko. Sporofyty různých rodů se výrazně liší a představují největší nižší rostliny, dosahující délky 60-100 m, s výrazným morfologickým členěním a složitou anatomickou stavbou. Ve většině případů jsou sporofyty řasy rozděleny na čepel ve tvaru listu, „kmen“ a rhizoidy, pomocí kterých je celá rostlina připevněna k podvodním kamenům a skalám. Na rozhraní čepele listu a stonku se nachází interkalární meristém, jehož činností listová čepel i stonek rostou. rod řasy, jejíž druhy jsou rozšířeny v severních mořích. 63
TŘÍDA HETEROGENERÁTNÍ Řád Laminaria Gametofyty se od sebe málo liší a jsou zastoupeny mikroskopickými, často na několik buněk redukovanými, vláknitými rostlinami nesoucími rozmnožovací orgány. Na samčích gametofytech se tvoří antheridia ve formě malých buněk, které vyvíjejí jednu spermii, na samičích gametofytech - oogonia, ve kterých se tvoří jedno vajíčko. Sporofyty různých rodů se výrazně liší a představují největší nižší rostliny, dosahující délky 60-100 m, s výrazným morfologickým členěním a složitou anatomickou stavbou. Ve většině případů jsou sporofyty řasy rozděleny na čepel ve tvaru listu, „kmen“ a rhizoidy, pomocí kterých je celá rostlina připevněna k podvodním kamenům a skalám. Na rozhraní čepele listu a stonku se nachází interkalární meristém, jehož činností listová čepel i stonek rostou. rod řasy, jejíž druhy jsou rozšířeny v severních mořích. 63
 TŘÍDA CYCLOSPORONAE Řád Fucus je charakterizován - - apikálním růstem thallusu Absence nepohlavního rozmnožování oogamním pohlavním procesem.Pohlavní orgány jsou umístěny v prohlubních thallusu - pojmů, neboli scafidia.Dochází k vegetativnímu rozmnožování, vedoucí v některých částech oceánu ke vzniku obrovských nahromadění Sargassum thalli (příkladem je Sargasové moře).
TŘÍDA CYCLOSPORONAE Řád Fucus je charakterizován - - apikálním růstem thallusu Absence nepohlavního rozmnožování oogamním pohlavním procesem.Pohlavní orgány jsou umístěny v prohlubních thallusu - pojmů, neboli scafidia.Dochází k vegetativnímu rozmnožování, vedoucí v některých částech oceánu ke vzniku obrovských nahromadění Sargassum thalli (příkladem je Sargasové moře).
 rod Sargassum komplexní morfologické dělení thallusu je rozšířeno na jižní polokouli, zejména v teplých mořích. Stonek je přichycen k substrátu podrážkou, větví, nese zploštělé listovité útvary, kulovité vzduchové bubliny na zvláštních stopkách, rozvětvené plodné větve 71
rod Sargassum komplexní morfologické dělení thallusu je rozšířeno na jižní polokouli, zejména v teplých mořích. Stonek je přichycen k substrátu podrážkou, větví, nese zploštělé listovité útvary, kulovité vzduchové bubliny na zvláštních stopkách, rozvětvené plodné větve 71



 Hnědé řasy jsou přirozenou skupinou, která se svou morfologií dobře odlišuje od ostatních oddělení řas. Jak však již bylo zmíněno, podle řady charakteristik (přítomnost chlorofylu „a“ a „c“, nepřítomnost chlorofylu „b“, umístění thylakoidů v chloroplastech – trithylakoidní lamely, podle podobných skladovacích produktů – řasa , chrysolamin, podle struktury monadických buněk s heterokontem a heteromorfními bičíky) hnědé řasy vykazují podobnosti se zlatými, žlutozelenými, rozsivek a pyrofytními řasami. Na tomto základě někteří autoři zařazují hnědé a další uvedené skupiny řas do třídy tříd v rámci jedné velké divize Credoshore. bу1 a. Existence monadických buněk v životním cyklu hnědých řas umožňuje, aby byly stejně jako ostatní uvedené skupiny odvozeny od některých primárních fotosyntetických bičíků s převahou hnědých pigmentů. Tito bičíkovci se vyvinuli několika způsoby, z nichž jeden vedl ke vzniku hnědých řas. Hnědé řasy však mají s bičíkovci vzdálenější spojení než zlaté řasy, heterobičíkovce, rozsivky, pyrofyty a zelené řasy, které jsou si v řadě vlastností podobné, stejně jako zelené řasy, neboť zde neexistují žádné přímé přechodné formy. Mezi hnědými řasami neexistuje konzistentní vývoj typů organizace stélků od monadických po vláknité a lamelární, jak lze snadno pozorovat u zelených, žlutozelených, zlatých a pyrofytických řas. Jak již bylo uvedeno, mezi hnědými řasami jsou zastoupeny pouze nejvyšší stupně morfologické diferenciace stélky - heterotrichální a lamelární. Snad jednodušší formy (monáda, kokoid, jednoduchý vláknitý) se během dlouhé evoluce prováděné hnědými řasami - starověkou již známou skupinou, zcela ztratily; ze silurských a devonských nalezišť. Neexistuje jednotný pohled na související vztahy zakázek v rámci oddělení hnědých řas a jejich umístění v systému. Podle jednoho ze schémat fylogeneze hnědých řas, konstruovaného s přihlédnutím k podobnosti struktury stélky v nejranějších fázích vývoje, jsou hnědé řasy difyletickou skupinou: jedna linie evoluce se spojuje (z těch, které jsou uvedeny v prezentaci ) Sphacelariaceae, Dictyotaceae a Fucus, ostatní - ektocarpaceae, Cutleriaceae a Laminariaceae. Oba 219" 75
Hnědé řasy jsou přirozenou skupinou, která se svou morfologií dobře odlišuje od ostatních oddělení řas. Jak však již bylo zmíněno, podle řady charakteristik (přítomnost chlorofylu „a“ a „c“, nepřítomnost chlorofylu „b“, umístění thylakoidů v chloroplastech – trithylakoidní lamely, podle podobných skladovacích produktů – řasa , chrysolamin, podle struktury monadických buněk s heterokontem a heteromorfními bičíky) hnědé řasy vykazují podobnosti se zlatými, žlutozelenými, rozsivek a pyrofytními řasami. Na tomto základě někteří autoři zařazují hnědé a další uvedené skupiny řas do třídy tříd v rámci jedné velké divize Credoshore. bу1 a. Existence monadických buněk v životním cyklu hnědých řas umožňuje, aby byly stejně jako ostatní uvedené skupiny odvozeny od některých primárních fotosyntetických bičíků s převahou hnědých pigmentů. Tito bičíkovci se vyvinuli několika způsoby, z nichž jeden vedl ke vzniku hnědých řas. Hnědé řasy však mají s bičíkovci vzdálenější spojení než zlaté řasy, heterobičíkovce, rozsivky, pyrofyty a zelené řasy, které jsou si v řadě vlastností podobné, stejně jako zelené řasy, neboť zde neexistují žádné přímé přechodné formy. Mezi hnědými řasami neexistuje konzistentní vývoj typů organizace stélků od monadických po vláknité a lamelární, jak lze snadno pozorovat u zelených, žlutozelených, zlatých a pyrofytických řas. Jak již bylo uvedeno, mezi hnědými řasami jsou zastoupeny pouze nejvyšší stupně morfologické diferenciace stélky - heterotrichální a lamelární. Snad jednodušší formy (monáda, kokoid, jednoduchý vláknitý) se během dlouhé evoluce prováděné hnědými řasami - starověkou již známou skupinou, zcela ztratily; ze silurských a devonských nalezišť. Neexistuje jednotný pohled na související vztahy zakázek v rámci oddělení hnědých řas a jejich umístění v systému. Podle jednoho ze schémat fylogeneze hnědých řas, konstruovaného s přihlédnutím k podobnosti struktury stélky v nejranějších fázích vývoje, jsou hnědé řasy difyletickou skupinou: jedna linie evoluce se spojuje (z těch, které jsou uvedeny v prezentaci ) Sphacelariaceae, Dictyotaceae a Fucus, ostatní - ektocarpaceae, Cutleriaceae a Laminariaceae. Oba 219" 75
 Skupiny řádů se liší i způsobem růstu thalli: první se vyznačuje apikálním růstem, druhý interkalárním růstem. Kilin (N. Ku 1sh), jehož systém je v současnosti nejrozšířenější, odvozuje od společného předka hnědých řas tři evoluční linie, které prezentuje jako třídy izogenerátní, heterogenerátní a cyklosporózní; jsou založeny na rozdílech v životních cyklech a morfologické struktuře řas. Velké hnědé řasy jsou široce používány v lidském zemědělství. Extrahuje se z nich algin – přilnavá látka používaná v textilním, potravinářském a řadě dalších odvětví. V pobřežních zemích se emise z mořských řas, bohaté na draslík a dusík, používají jako hnojivo a také jako krmivo pro dobytek. Některé, jako je kelp (mořské řasy), jsou jedlé. 76
Skupiny řádů se liší i způsobem růstu thalli: první se vyznačuje apikálním růstem, druhý interkalárním růstem. Kilin (N. Ku 1sh), jehož systém je v současnosti nejrozšířenější, odvozuje od společného předka hnědých řas tři evoluční linie, které prezentuje jako třídy izogenerátní, heterogenerátní a cyklosporózní; jsou založeny na rozdílech v životních cyklech a morfologické struktuře řas. Velké hnědé řasy jsou široce používány v lidském zemědělství. Extrahuje se z nich algin – přilnavá látka používaná v textilním, potravinářském a řadě dalších odvětví. V pobřežních zemích se emise z mořských řas, bohaté na draslík a dusík, používají jako hnojivo a také jako krmivo pro dobytek. Některé, jako je kelp (mořské řasy), jsou jedlé. 76
Ve velkých hloubkách, až 250 metrů, rostou červené řasy, jinak nazývaný šarlatový. Různě zbarvené šarlatové ryby vytvářejí v kombinaci s korály a zářivými rybami jedinečnou krásu podmořského světa. Jedná se převážně o velké řasy, ale například mikroskopické červené řasy patří také do třídy Banguiaceae.
Proč mohou červené řasy růst v tak významných hloubkách? Tato otázka byla položena v Jednotné státní zkoušce z biologie. Červený pigment umožňuje řasám růst ve velkých hloubkách fykoerythrin. Díky ní při fotosyntéze pohlcují červené řasy zelené, modré, modrofialové paprsky spektra. Právě tyto paprsky, na rozdíl od červených, jsou schopny proniknout hluboko do vodního sloupce.
Červené řasy se vyznačují asexuálním rozmnožováním spor, stejně jako pohlavním rozmnožováním (oogamie); někdy se vyskytuje i vegetativní rozmnožování částmi stélku.
Mezi červené řasy patří phyllophora, porphyra, gracillaria, ptilote, chondrius a celkem jich je asi pět tisíc druhů.
Nachový- plochá a tenká oválná deska do průměru půl metru. Vyznačuje se pouze pohlavním rozmnožováním. Mužské reprodukční buňky nemají bičíky (spermie). Je to pochopitelné, protože ve velkých hloubkách pod vodním sloupcem je obtížné se pohybovat pomocí bičíku.
Význam řas
Místo v potravních řetězcích, dopad na přírodu
1. Obrovská masa řas vytváří fytoplankton a i v arktických mořích připadá 20-30 milionů jedinců na 1 kubický metr vody. Jedná se o primární produkty, které tvoří základ potravního řetězce.
2. Fytoplankton slouží jako potrava pro zooplankton (sekundární produkty), které jedí velcí mořští obyvatelé, jako jsou velryby. Je zajímavé, že Thor Heyerdahl během expedice na raftu Kon-Tiki vyzkoušel planktonovou „polévku“ a zjistil, že je docela chutná a výživná.
3. Spodní řasy poskytují úkryt rybám a různým mořským živočichům a slouží jim také jako potrava. Například řasu s radostí pojídají mořští ježci.
4. Řasy nasycují oceány a atmosféru kyslíkem.
5. Když se však řasy (například Chlamydomonas) masivně množí během tzv. „rozkvětu“ vody, obsah kyslíku v ní klesá a voda se nasytí toxiny. Obyvatelé řek umírají na nedostatek kyslíku.
Význam pro lidi
1. Řasy se konzumují jako potrava v mnoha regionech po tisíce let. Obzvláště oblíbené jsou hnědé a červené řasy jako chaluha, undaria, porfyra, hijiki (a sargassum obecně).
2. Červené řasy jsou zdrojem jódu, zejména některé druhy na něj bohaté.
3. Červené řasy jsou také zdrojem agar-agaru, gelotvorné látky, která se používá v cukrářském průmyslu, ke kultivaci bakterií atp.
4. Pomocí řas se čistí odpadní voda od fluoru, dusíku atd. a vzduch od oxidu uhličitého (to se podařilo Chlamydomonas, Chlorella, Euglena).
5. Řasy jsou potravinářské přídatné látky: spirulina, chaluha, fucus, ulva, chlorella a další.