Příklady alelických a nealelických genů. Interakce alelických genů
Lidská genetika se základy obecné genetiky [Výuka] Kurchanov Nikolay Anatoljevič
4.3. Genová interakce
4.3. Genová interakce
Mnoho genů funguje v těle současně. V procesech implementace genetické informace do znaku jsou možné četné „body“ interakce mezi různými geny na úrovni biochemických reakcí. Takové interakce nevyhnutelně ovlivňují tvorbu fenotypu.
Alelické geny určují alternativní vlastnosti, protože leží v homologních lokusech homologních chromozomů. Mezi alelami heterozygota jsou možné určité interakce, které jsou základem projevu znaku ve fenotypu. Jsou známy tři hlavní formy interalelických interakcí.
Úplná dominance - u heterozygota potlačuje jedna alela (dominantní) projev druhé alely (recesivní).
Neúplná dominance – u heterozygota dochází k intermediárnímu projevu vlastností alel.
Kodominance je nezávislým projevem alel u heterozygota.
V některých případech se podařilo rozluštit mechanismus interakce alel. Jejich vztah nejlépe ilustrují enzymové proteiny. Data z biochemické genetiky ukazují, že fenomén dominance je často spojen s aktivitou konkrétního enzymu.
Méně jasný je fenomén monogenní heterózy, kdy heterozygot vykazuje znak pro dominantní alelu silněji než homozygot. Heteróza- fenomén nadřazenosti kříženců nad oběma rodičovskými formami. Fenomén heterózy byl intenzivně studován pro jeho důležitou roli při selekci. Tento fenomén je velmi složitý a nejednoznačný. Pojem „nadřazenost“ je nejednoznačný, protože heteróza se projevuje na reprodukční, somatické a adaptivní úrovni. Projevy na různých úrovních mohou být opačného směru (např. „zlepšení“ somatických ukazatelů může být doprovázeno „zhoršením“ adaptivních). Samotné pojmy „zlepšení“ a „zhoršení“ jsou nejednoznačné, protože nemají jasná kritéria a jejich aplikace je extrémně subjektivní. Existuje několik teorií vysvětlujících povahu heterózy.
Fenomén monogenní heterózy, kdy je ukázána závislost znaku na jednom alelickém páru, je pouze jedním aspektem teorie heterózy. Na návrh F. Dobzhanského byl tento fenomén tzv převaha. Pro evoluční teorii má velký význam, protože ukazuje výhodu heterozygotů v populacích. Tento koncept je však aplikovatelný spíše na adaptivní úroveň, takže dokud nebudou vyjasněny genetické mechanismy overdominance, je předčasné jej považovat za zvláštní typ interalelických interakcí.
V živých organismech často interagují nejen alely jednoho genu, ale také alely různých genů, což poskytuje širokou škálu možností segregace. Existují tři hlavní typy interakce mezi nealelickými geny.
Komplementarita - interakce různých dominantních alel způsobuje objevení se nového znaku.
Geny, které řídí různá stádia stejné metabolické dráhy, obvykle interagují podle typu komplementarity. U některých morfologických znaků je však biochemický mechanismus implementace neznámý.
Epistáza – jeden gen potlačuje expresi jiného, nealelického genu.
Geny, které potlačují působení jiných genů, se nazývají epistatické (neboli supresorové geny). Existují dvě možné možnosti epistázy: dominantní epistáze– epistatický gen je dominantní ve svém alelickém páru a recesivní epistáze– epistatický gen je ve svém alelickém páru recesivní.
Polymerismus je jednoznačné působení nealelických genů. Polymerismus je spojen s kontrolou vlastnosti několika nealelickými geny. Polygenní kontrola je v genetice velmi rozšířená. Polymerní geny jsou obvykle označeny stejnými písmeny s dolním indexem - A 1, A 2, A 3 atd.
Polymeria se také vyskytuje ve dvou odrůdách. Na kumulativní polymer intenzita znaku je úměrná počtu dominantních alel mezi polymerními geny a kdy nekumulativní polymer různé polymerní geny se navzájem duplikují, a aby se znak projevil, stačí přítomnost jedné z dominantních alel.
Četné případy interakce genů zaplňují většinu knih o genetických problémech. V typickém případě při křížení diheterozygotů má interakce genů za následek vznik velmi odlišných poměrů fenotypových tříd v generacích - 9: 3: 4; 9:7; 13:3; 12:3:1; 15:1 a další. Genetická analýza ukazuje, že jsou to všechny modifikace klasického mendelovského dihybridního vzorce štěpení 9:3:3:1. Řešení velkého množství problémů v genetice je nezbytným krokem v přípravě studentů studujících genetiku.
Fráze „genová interakce“ je poněkud svévolná, protože obvykle neinteragují samotné geny, ale jejich produkty. Nemůžeme však souhlasit s termínem „interakce vysoušečů vlasů“, který přesně neodráží význam tohoto jevu. Podle mého názoru je lepší v naučné literatuře opustit tradiční termín „genová interakce“ (alelický i nealelický).
Tento text je úvodním fragmentem. Z knihy Genetika barev psů od Robinsona RoyeInterakce lokusů A a E Význam těchto dvou sérií alel bude ukázán v této části. Obě řady kontrolují distribuci černých a žlutých pigmentů v srsti většiny plemen psů. V interakci tvoří tyto lokusy mnoho dobře známých
Z knihy Mikrobiologie: poznámky k přednášce autor Tkačenko Ksenia Viktorovna2. Interakce virů s hostitelskou buňkou Interakce probíhá v jediném biologickém systému na genetické úrovni.Existují čtyři typy interakce: 1) produktivní virová infekce (interakce vedoucí k reprodukci viru, a
Z knihy Mravenec, rodina, kolonie autor Zacharov Anatolij AlexandrovičINTERAKCE MRAVENCŮ V RODINĚ Rodina mravenců je sdružení stovek, tisíců a někdy i milionů jedinců, jejichž úsilím si staví hnízdo, krmí četné potomstvo, chrání mraveniště a jeho krmnou plochu před zásahy agresivních
Z knihy Evoluce autor Jenkins MortonGENOVÝ DRIFT Tento koncept se někdy nazývá „Sewell-Wrightův efekt“ podle dvou populačních genetiků, kteří jej navrhli. Poté, co Mendel dokázal, že geny jsou jednotkami dědičnosti, a Hardy a Weinberg prokázali mechanismus jejich chování,
Z knihy Lidská rasa od Barnetta AnthonyhoInterakce dědičnosti a prostředí Někdy se lidé ptají: co je důležitější - dědičnost nebo prostředí? Na tuto otázku není tak snadné odpovědět. Pokud tím myslíme co? má největší dopad, pak i tehdy bychom se měli omezit na zvláštní případy.
Z knihy Základy psychofyziologie autor Alexandrov Jurij7. INTERAKCE SMYSLOVÝCH SYSTÉMŮ K interakci senzorických systémů dochází na spinální, retikulární, thalamické a kortikální úrovni. Integrace signálů v retikulární formaci je obzvláště široká. K integraci signálů vyššího řádu dochází v mozkové kůře. V
Z knihy Embrya, geny a evoluce od Raffa Rudolfa AInterakce a integrace Návštěvníci muzea dnes považují vystavené namontované kostry obrovských vyhynulých zvířat za samozřejmost. Ne vždy tomu tak však bylo. Vědecké informace o tak působivých formách, jako jsou mamuti a
Z knihy Teorie adekvátní výživy a trofologie [tabulky v textu] autor Z knihy Stop, kdo vede? [Biologie chování lidí a jiných zvířat] autor Žukov. Dmitrij AnatoljevičInterakce endokrinních systémů: přímá komunikace, zpětná vazba, synergismus, permisivní působení, antagonismus Jak již bylo řečeno, jednotlivé žlázy s vnitřní sekrecí a jejich hormony se vzájemně ovlivňují a zajišťují funkční jednotu těla. Mezi
Z knihy Teorie adekvátní výživy a trofologie [tabulky s obrázky] autor Ugolev Alexandr Michajlovič9.7. Závěrečné poznámky (interakce v biocenózách) Uvažujme o biochemických interakcích živých organismů v přírodních podmínkách, které jsou mimořádně rozšířené a staly se předmětem speciální vědy - alelopatie (viz kap. 1). Příklady takových
Z knihy Fenetika [Evoluce, populace, rys] autor Jablokov Alexej VladimirovičInterakce evolučních sil - mechanismus evoluce Spouštěcí mechanismus evoluce funguje jako výsledek společného působení evolučních faktorů uvnitř populace jako evoluční jednotky. Každá populace každého druhu je vystavena tlakům toho či onoho druhu.
Z knihy Evoluce [Klasické myšlenky ve světle nových objevů] autorGenová duplikace MULTIFUNKČNÍ GENY – ZÁKLAD EVOLUČNÍCH INOVACÍ Myšlenku, že genová duplikace slouží jako nejdůležitější zdroj evolučních inovací, vyslovil již ve 30. letech 20. století vynikající biolog John Haldane (Haldane, 1933). Dnes neexistuje
Z knihy Geny a vývoj těla autor Neyfakh Alexandr Alexandrovič1. Genové promotory V této části stručně pohovoříme o tom, které nukleotidové sekvence sousedící s geny a někdy uvnitř genu jsou zodpovědné za proces transkripce. U prokaryot tyto oblasti, na které se molekula RNA polymerázy váže a odkud
Z knihy Evoluce člověka. Kniha 1. Opice, kosti a geny autor Markov Alexandr VladimirovičZměny v genové aktivitě K evoluci zvířat obecně a primátů zvláště nedochází ani tak v důsledku změn ve struktuře genů kódujících proteiny, jako spíše v důsledku změn jejich aktivity. Malá změna v horních patrech organizovaná hierarchicky
Z knihy Genetika člověka se základy obecné genetiky [Výukový program] autor Kurčanov Nikolaj Anatolievič4.4. Interakce genotypu a prostředí Povaha projevu působení genů je mnohem složitější než u výše popsaných možností. Při zvažování vlivu genů a jejich alel je třeba vzít v úvahu vliv vnějšího prostředí na projevy znaků a také modifikační vliv jiných
Z autorovy knihy7.1. Izolace genů Lze použít několik způsobů izolace genů. Každý z nich má své výhody a nevýhody Chemická syntéza genů, tedy syntéza nukleotidů s danou sekvencí odpovídající jednomu genu, byla poprvé provedena v r.
Interakce mezi alelickými geny probíhá ve třech formách: úplná dominance, neúplná dominance a nezávislá manifestace (kodominance).
Úplná dominance – když jedna dominantní alela zcela potlačí projev alely recesivní, např. žlutá barva hrášku je dominantní nad barvou zelenou.
Neúplná dominance je pozorována, když jeden gen z páru alel neposkytuje tvorbu svého proteinového produktu dostatečného pro normální projev znaku. Při této formě genové interakce se všichni heterozygoti a homozygoti navzájem významně liší fenotypem. Příkladem štěpení s neúplnou dominancí je dědičnost barvy květů Night Beauty.
Při křížení rostlin s červenými květy (AA) a rostlin s bílými květy (aa) mají hybridy F1 růžové květy (Aa). Dochází tak k neúplné dominanci; v F2 je rozdělení 1:2:1 jak ve fenotypu, tak v genotypu.
Kromě úplné a neúplné dominance jsou známy případy absence dominantně recesivních vztahů či kodominance. Když je v heterozygotních organismech kodominantní, každý z alelických genů způsobí vytvoření vlastnosti, kterou řídí ve fenotypu.
Příkladem této formy interakce alel je dědičnost lidských krevních skupin podle systému AB0, určená genem I. Existují tři alely tohoto genu Io, Ia, Ib, které určují antigeny krevních skupin. Dědičnost krevních skupin ilustruje fenomén mnohočetného alelismu: v genofondu lidských populací existuje gen I ve formě tří různých alel, které se u jednotlivých jedinců kombinují pouze v párech. Před tímto příkladem jsme hovořili o genech, které existují pouze ve dvou různých alelických formách. Mnoho genů se však skládá ze stovek párů nukleotidů, takže mutace se mohou vyskytovat v mnoha oblastech genu a dát vznik mnoha různým alelickým formám. Protože každý z homologních chromozomů má jeden alelický gen, pak samozřejmě diploidní organismus nemá v populačním genofondu více než dvě z řady alel.
30. Nealelické genové interakce
Nealelické geny- jedná se o geny umístěné v různých částech chromozomů a kódující různé proteiny. Nealelické geny mohou také vzájemně interagovat.
V tomto případě buď jeden gen určuje vývoj více znaků, nebo se naopak jeden znak projeví pod vlivem kombinace více genů. Existují tři formy a interakce nealelických genů:
komplementarita;
polymerismus.
Komplementární(dodatečné) působení genu je typem interakce nealelických genů, jejichž dominantní alely po spojení v genotyp určují nový fenotypový projev znaků. V tomto případě může dojít k štěpení F2 hybridů podle fenotypu v poměrech 9:6:1, 9:3:4, 9:7, někdy 9:3:3:1. Příkladem komplementarity je dědičnost tvaru plodu dýně. Přítomnost dominantních genů A nebo B v genotypu určuje kulovitý tvar plodu a protáhlý tvar recesivních genů. Pokud genotyp obsahuje oba dominantní geny A i B, bude tvar plodu diskoidní. Při křížení čistých linií s odrůdami, které mají kulovitý tvar plodů, budou mít v první hybridní generaci F1 všechny plody diskovitý tvar a v generaci F2 dojde k rozštěpení fenotypu: z každých 16 rostlin 9 budou mít plody diskovitého tvaru, 6 - kulovité a 1 - protáhlé .
Epistáze- interakce nealelických genů, při které je jeden z nich potlačen druhým. Supresivní gen se nazývá epistatický, suprimovaný gen se nazývá hypostatický. Pokud epistatický gen nemá svůj vlastní fenotypový projev, pak se nazývá inhibitor a označuje se písmenem I. Epistatická interakce nealelických genů může být dominantní a recesivní. U dominantní epistázy je manifestace hypostatického genu (B, b) potlačena dominantním epistatickým genem (I > B, b). Štěpení fenotypu při dominantní epistáze může probíhat v poměru 12:3:1, 13:3, 7:6:3. Recesivní epistáze je potlačení alel hypostatického genu (i > B, b) recesivní alelou epistatického genu. Fenotypové štěpení může probíhat v poměru 9:3:4, 9:7, 13:3.
Polymerismus- interakce nealelických více genů, které jednoznačně ovlivňují vývoj stejného znaku; Stupeň projevu znaku závisí na počtu genů. Polymerní geny jsou označeny stejnými písmeny a alely stejného lokusu mají stejný dolní index.
Polymerní interakce nealelických genů může být kumulativní a nekumulativní. Při kumulativní (akumulativní) polymeraci závisí stupeň projevu znaku na sumativním působení genů. Čím více dominantních genových alel, tím výraznější je konkrétní znak. Štěpení F2 ve fenotypu probíhá v poměru 1:4:6:4:1.
Při nekumulativní polymeraci se znak projevuje v přítomnosti alespoň jedné z dominantních alel polymerních genů. Počet dominantních alel neovlivňuje stupeň exprese znaku. K fenotypové segregaci dochází v poměru 15:1.
Příklad: barva kůže u lidí, která závisí na čtyřech genech.
Alelické geny jsou geny umístěné v identických oblastech homologních chromozomů a řídící vývoj variací jednoho znaku.
Nealelické geny se nacházejí v různých částech homologních chromozomů a řídí vývoj různých znaků.
Koncept působení genů.
Gen je úsek molekuly DNA nebo RNA, který kóduje sekvenci nukleotidů v tRNA a rRNA nebo sekvenci aminokyselin v polypeptidu.
Charakteristika působení genu:
Gen je diskrétní
Gen je specifický – každý gen je zodpovědný za syntézu přísně specifické látky
Gen působí postupně
Pleiotropní efekt - 1 gen působí na změnu nebo projev několika příznaků (1910 Plate) fenylketonurie, Marfanův syndrom
Působení polymeru – pro expresi vlastnosti je potřeba několik genů (1908 Nilsson-Ehle)
Geny se vzájemně ovlivňují prostřednictvím jimi určených proteinových produktů
Genová exprese je ovlivněna faktory prostředí
Vyjmenujte typy interakcí mezi alelickými a nealelickými geny.
Mezi alelami:
Úplná nadvláda
Neúplná dominance
Kodominance
Převaha
Mezi nealelickými: (znak nebo vlastnosti jsou určeny dvěma nebo více nealelickými geny, které se vzájemně ovlivňují. I když zde je interakce podmíněná, protože neinteragují geny, ale produkty jimi řízené. V v tomto případě jde o odchylku od mendělejevských zákonů segregace).
Polymerismus
Komplementarita
Podstata naprosté dominance. Příklady.
Úplná dominance je typ interakce alelických genů, kdy dominantní gen (A) zcela potlačuje působení recesivního genu (a) (pihy)
Neúplná dominance. Příklady.
Neúplná dominance je typ interakce alelických genů, kdy dominantní alela zcela nepotlačuje působení recesivní alely a tvoří znak se středním stupněm degenerace (barva očí, tvar vlasů)
Převaha jako základ heterózy. Příklady.
Overdominance je typ interakce alelických genů, kdy gen v heterozygotním stavu má větší fenotypový projev znaku než homozygotní.
Srpkovitá anémie. A – hemoglobinA a – hemoglobinS. AA – 100% normální červené krvinky, náchylnější k malárii; aa – 100% mutovaný (zemře), Aa – 50% mutovaný, prakticky není náchylný k malárii, protože už ohromen
Kodominance a její podstata. Příklady.
Kodominance je typ interakce alelických genů, kdy se na určení znaku podílí několik alel genu a vytvoří se znak nový. Jeden alelický gen doplňuje působení jiného alelického genu, nový znak se liší od rodičovských (krevní skupina ABO).
Fenomén nezávislého projevu obou alel ve fenotypu heterozygota, jinými slovy absence dominantně recesivních vztahů mezi alelami. Nejznámějším příkladem je interakce alel, které určují čtvrtou lidskou krevní skupinu (AB). Je známa vícenásobná série, která se skládá ze tří alel genu I, který určuje znamení krevní skupiny člověka. Gen I je zodpovědný za syntézu enzymů, které připojují určité polysacharidy k proteinům umístěným na povrchu červených krvinek. (Tyto polysacharidy na povrchu červených krvinek určují specifičnost krevních skupin.) Alely 1 A a 1 B kódují dva různé enzymy; alela 1° nic nekóduje. V tomto případě je alela 1° recesivní ve vztahu k 1 A i I B a mezi posledními dvěma neexistuje žádný dominantně recesivní vztah. Lidé se čtvrtou krevní skupinou nesou ve svém genotypu dvě alely: 1 A a 1 B. Vzhledem k tomu, že mezi těmito dvěma alelami neexistuje žádný dominantně-recesivní vztah, jsou v těle takových lidí syntetizovány oba enzymy a vzniká odpovídající fenotyp – čtvrtá krevní skupina.
Naprostá dominance. Na příkladu pokusů G. Mendela s hrachem je zřejmé, že jeden gen může zcela potlačit projev jiného alelického genu.
Neúplná dominance. Při neúplné dominanci se znak objevuje i v první generaci mezi rodičovskými formami. Pokud spářite karakulské ovce s dlouhými ušima s bezušními, pak v F1 budou všichni potomci krátkouší. Rozdělení v F2 podle fenotypu a genotypu se shoduje (1:2:1).
Kodominance. Oba alelické geny se objevují ve fenotypu a nevykazují mezi sebou ani dominanci ani recesivitu. U lidí jsou známy krevní systémy ABO, které určují erytrocytární antigeny. Alely A a B interagují podle typu kodominance.
V letech 1940 až 1970 bylo u skotu objeveno 12 genetických systémů krevních skupin. Rozlišují více než 100 krevních antigenů, definujících 369 fenoskupin, které mohou činit asi dva biliony různých kombinací – sérologických typů. fenoskupina- skupina antigenů (někdy jeden antigen), které jsou fenotypovou expresí souboru jednotlivých genů jednoho lokusu. Krevní skupiny, které jsou založeny na individuálních charakteristikách antigenních vlastností červených krvinek, jsou dědičné a po celý život zvířete se nemění. Dědí se jednotlivě nebo komplexně, a proto mohou sloužit jako vhodný genetický model při řešení mnoha teoretických a praktických problémů selekce, protože převážný počet známých alelických erytocytárních faktorů se dědí podle typu kodominance.
Převaha. Vyznačuje se tím, že u heterozygotních jedinců je znak výraznější než u homozygotních dominantních jedinců, to znamená, že kříženec je ve vývoji znaku nadřazen oběma rodičům. Tento jev se nazývá heterozas (Aa>AA a aa). Jasným příkladem převahy je lidská srpkovitá anémie. SS homozygoti mají některé červené krvinky ve tvaru srpu a šípu a jejich transport kyslíku v červených krvinkách je narušen. Lidé trpí akutní anémií a většinou umírají v raném věku. Heterozygoti Ss jsou odolní vůči tropické malárii, homozygoti Ss rozvíjejí tropickou malárii.
Mnohonásobný alelismus. Uvažovali jsme především o interakci dvou alelických genů, z nichž každý se může v důsledku mutace změnit a vznikne nový alelický gen. Tento jev se nazývá „mnohonásobný alelismus“. U lidí je příkladem mnohočetného alelismu krevní systém ABO, který má 3 alelické geny (A, B, O) a podle toho 6 genotypů a 4 fenotypy.
U králíků byly identifikovány následující alely, které ovlivňují barvu srsti: C - černá, cсch - činčila, ch - himálajská, ca - albino. Alely jsou uvedeny v sestupném pořadí podle dominance (C>cch>ch>ca). Alela C je dominantní nad všemi následujícími, ca je recesivní vůči všem předchozím (obr. 2.5).
Pleiotropní účinek genu. Gen může ovlivnit vývoj dvou nebo více vlastností, to znamená, že jsou pozorovány vícenásobné účinky genu. Tento typ interakce byl objeven při chovu šedých (shirazi) a černých (arabi) karakulských ovcí. Jehňata homozygotní pro dominantní alelu genu C, která určuje šedou barvu (Shirazi). při přechodu na pastvu umírají v důsledku nedostatečného rozvoje parasympatického nervového systému bachoru. Je to způsobeno působením dvou dominantních genů, které mají dvojí (pleiotropní) účinek: způsobují šedé zbarvení a v homozygotním stavu poruchu nervového systému.
Akademik D. K. Belyaev a jeho kolegové zjistili, že u norků jsou některé mutace spojené se změnami barvy srsti recesivní a v důsledku pleiotropního efektu se snižuje plodnost a životaschopnost zvířat.
Smrtelné a pololegální geny. Geny způsobují složité fyziologické změny během vývoje organismu. Mohou snížit vitalitu těla, a to až k smrti.
Jeden z důvodů, který mění štípání v poměru 3:1. je rozdílná životaschopnost homozygotů v F2. Ve 20. století skutečnosti byly objeveny, když došlo k rozdělení fenotypu 2:1. Při studiu dědičnosti barvy u žlutých a černých myší bylo tedy pozorováno rozdělení barvy na 2 části žluté a 1 část černé myši. V tomto případě některé žluté myši uhynuly během embryonálního období. Stejný gen v dominantním heterozygotním stavu způsobil žluté zbarvení myší a v homozygotním stavu vykazoval letální účinek. Gen černé barvy je alelický a recesivní.
Ve 30. letech XX století. U stříbrnočerné lišky bylo objeveno nové zbarvení, které se nazývá platina. Při křížení platinových lišek mezi sebou měli jedinci platinové i stříbrnočerné zbarvení v poměru 2:1. Při provádění analytického křížení se ukázalo, že dominantní je gen platinové barvy. Bylo navrženo, že homozygotní platinové lišky (AA) umírají během embryonálního období. Pitvy březích samic tento předpoklad potvrdily.
Recesivní letální geny působí v homozygotním stavu a dominantní také působí v heterozygotním stavu. Pokud gen nezpůsobí smrt organismu, ale výrazně sníží životaschopnost, pak se nazývá semiletální.
Smrtelné a pololegální geny jsou nejčastěji recesivní. Letální geny způsobují smrt organismu v embryonálním nebo postembryonálním období. Například recesivní legální geny způsobují absenci srsti u novorozených telat, králíků a zvířat jiných druhů. Brzy po narození tato zvířata umírají.
Pololegální geny mohou u zvířat způsobit deformace: zkrácení osového skeletu; deformace hlavy a končetin; porušení fyziologických funkcí těla. Chovná zvířata s letálními a semiletálními geny musí být utracena. Zejména je nutné kontrolovat chovné plemeníky, od kterých je umělou inseminací získáváno velké množství potomků.
Legální geny se mohou volně šířit z jedné země do druhé. Plemenný býk nizozemského plemene Prince Adolf, přenašeč genetické anomálie, byl tedy do Švédska přivezen z Nizozemska v roce 1902. O 26 let později vešlo ve známost, že býk je nositelem smrtícího recesivního genu pro bezsrstá telata. Jeho dcery, synové a vnuci byli hojně využíváni k chovatelským účelům a během poměrně krátké doby byla řada zvířat příbuzná s princem Adolfem. V některých stádech se asi 6 % telat narodilo bez srsti a zemřelo během několika minut po narození.
V poslední době se v zahraniční i domácí literatuře objevují zprávy o genetických anomáliích (defektech) způsobujících obrovské ekonomické škody v chovu hospodářských zvířat (P. D. Smith, L. F. Novíková). Takže například defekty BLAD (Bovine Leukocyte Adhesion Deficiency) nebo adheze (shlukování) leukocytů u skotu a CVM (Complex Vertebral Malformation) nebo komplexní deformace páteře mají recesivní typ dědičnosti a nejsou vyjádřeny v heterozygotní stav (obr. 2.6) . Tyto genetické vady se rozšířily v linii holštýnského skotu Montvik Chieftain 95679, která je široce používána v Ruské federaci. Jeden z nejznámějších býků této linie, Osbordale Ivanhoe 1189870, byl nositelem genetické vady B LAD. a jeho vnuk Carlin-M Ivanhoe Bell 1667366 je nositelem genetických defektů BLAD i CVM.
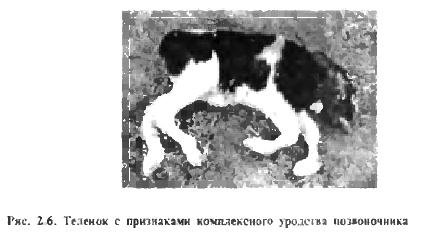
V průměru 2–5 % novorozených zvířat mnoha druhů má dědičné anomálie. Letální a semi-legální geny jsou jednou z příčin embryonální úmrtnosti a vrozených anomálií. Pro každý typ hospodářského zvířete bylo identifikováno několik desítek takových genů a byla uvedena mezinárodní klasifikace anomálií. Dědičný charakter mrtvorozených dětí, potratů a deformací dokládá skutečnost, že jejich procento se výrazně zvyšuje příbuzenskou plemenitbou.
Nejběžnější dědičné anomálie u skotu jsou: mopsovité; svalová kontraktura; absence předních a zadních končetin; hydrops plodu a mozku; kožní defekty: bezsrstost; zkrácení čelisti; pupeční kýla atd.
Shrneme-li data různých autorů, můžeme říci, že u skotu existuje více než 40 homozygotních letalit, deformací a karyotypových anomálií – dědičných sňatků. Bylo identifikováno 23 vrozených vad, které ovlivňují reprodukční funkci a plodnost krav.
U prasat bylo popsáno 33 genetických abnormalit. Nejběžnější jsou: cerebrální kýla; paralýza zadních končetin; nepřítomnost řitního otvoru; rozštěp patra; tlustonohý; zakřivení a tuhost končetin; kapání mozku; hemofilie.
U ovcí jsou nejvýraznější kraniofaciální vady: zkrácení nebo absence dolní čelisti; vypadávání vlasů, plešatost; nanismus; rozštěp patra aj. Celkem 43 anomálií.
Mezinárodní seznam právních vad obsahuje 45 anomálií u kuřat; 6 - u krůt; 3 - v kachnách; Z dědičných anomálií je 10 u koní.
Na dědičný původ anomálie naznačují následující skutečnosti: soulad zjištěné anomálie s popisem genetické vady, výskyt anomálie v důsledku příbuzenské plemenitby, projev obdobných anomálií u postranních příbuzných (sourozenci a pol. -sourozenci), příbuzní matky nebo otce.
Všechny případy projevů dědičných anomálií musí být zaznamenány do karet chovných zvířat, aby bylo možné v budoucnu provádět genetické analýzy. V této situaci hrají důležitou roli rodokmenové a objektivní chovné záznamy.Sběr dat pro posouzení genetické pohody chovných zvířat a přítomnosti letálních a semi-legálních genů by měl být prováděn v chovných podnicích a stanicích pro umělou inseminaci farem zvířat.
Interakce alelických genů a její typy
Při studiu zákonů G. Mendela jsme se již seznámili s hlavními typy interakce alelických genů. Na základě dříve prostudovaného materiálu vyplňte tabulku.
Stůl. Typy interakce mezi geny jednoho alelického páru
Typ interakce |
Povaha interakce v |
Vzorce pro fenotypové štěpení v F 2 a při testovacích křížení |
Příklady |
| Úplná nadvláda | Jedna alela (dominantní) zcela potlačuje účinek jiné (recesivní) alely |
Vysoká rostlina dominuje nanismu; kudrnaté vlasy - přes rovné vlasy; hnědé oči - nad modrými atd. |
|
| Neúplná dominance | Účinek každé alely není plně realizován, v důsledku čehož se objevuje přechodný znak |
Barvení květin noční krásy; srpkovitá anémie; cystonurie atd. |
|
| Kodominance | Účast obou alel na určování znaku |
IV krevní skupina (erytrocyty této krevní skupiny mají aglutinogeny typu A a typ V) |
Interakce nealelických genů
Na základě zákonů G. Mendela můžeme dospět k závěru, že mezi genem a znakem existuje poměrně silné spojení, že genotyp je tvořen souhrnem samostatně působících genů a fenotyp je mechanická kombinace individuálních rysů. Přímé a jednoznačné souvislosti mezi genem a znakem jsou však spíše výjimkou než pravidlem. Vývoj znaku v organismu je obvykle pod kontrolou mnoha genů a znak je výsledkem interakce nealelických genů.
Nejjednodušší případ interakce nealelických genů je, když je znak řízen dvěma páry alel. Podívejme se na několik příkladů takové interakce.
Komplementární genová interakce
Fenomén genové interakce, kdy je pro projev znaku nezbytná přítomnost dvou nealelických genů, se nazývá komplementarita(z lat. komplementum- sčítání), a geny nezbytné pro projev znaku jsou komplementární, nebo doplňkové.
Zvažme různé možnosti interakce komplementárních genů.
Alely druhého genu se objevují pouze v přítomnosti dominantní alely genuA. Zbarvení myší závisí v nejjednodušším případě na dvou genech. Myši s genotypem ahh nemají pigment a jsou bílé barvy. V přítomnosti dominantní alely A vyrobí se pigment a myš se nějak zbarví. Konkrétní barvu určuje druhý gen. Jeho dominantní alela S způsobuje šedou barvu myši a recesivní alelu S- Černá barva. Pokud tedy uvažujeme o homozygotních variantách, genotyp šedých myší je AASS, Černá - AAAss, bílá - aaSS nebo prdel. Při křížení šedé myši AASS s bílou prdel v první generaci vše dopadá podle Mendela: všichni hybridi jsou šedé barvy (jedná se o heterozygoty s genotypem AaSs). V F2, jak je snadné zkontrolovat, dostáváme 9/16 šedé myši, 3/16 černé a 4/16 bílé.

Aby se znak projevil, musí být v genotypu přítomny dominantní alely dvou různých genů. Příkladem takové komplementární genové interakce je dědičnost tvaru ovoce u dýní. V přítomnosti obou dominantních alel mají plody tvar disku, v přítomnosti jedné (jakékoli!) dominantní alely - kulovité a v nepřítomnosti dominantních alel - protáhlé.

Aby se znak projevil, musí být v genotypu přítomny dominantní alely dvou různých genů, ale každá dominantní alela v kombinaci s recesivními alelami druhého páru má nezávislý fenotypový projev. Například u kuřat je hrachovitý hřeben určen jedním dominantním genem, zatímco hřeben tvaru růže je určován jiným nealelickým, ale také dominantním genem. Když jsou tyto geny ve stejném genotypu, vyvine se hřeben ve tvaru ořechu. Pokud je organismus homozygotní pro oba recesivní geny, vyvine se jednoduchý hřeben ve tvaru listu. Při křížení dihybridů (vše s hřebenem ve tvaru ořechu) dochází ve druhé generaci ke štěpení v poměru 9:3:3:1. Zde však nelze nalézt nezávislou segregaci každé alely v poměru 3:1, protože ve všech případech shody v genotypu obou dominantních genů není detekován jejich přímý účinek.
O komplementární interakci nealelických genů tedy hovoříme v případě, kdy dva nealelické geny dávají nový znak, tedy dochází k nové formaci.
Epistatická genová interakce
Interakce genů, kdy jeden gen potlačuje působení jiného, nealelického k prvnímu, se nazývá epistáze(z řečtiny epi– výše a stáze- stand), a geny, které potlačují působení jiných genů se nazývají supresorové geny nebo inhibiční geny(z lat. inhiber– držet), popř epistatické geny. Gen, který je potlačován, se nazývá hypostatický.
Existují dominantní a recesivní epistáze.
Dominantní epistáze spojené s dominantním inhibitorovým genem. Například gen já určuje bílou barvu plodů dýně, je-li přítomen, vliv gen V neobjevuje se. S genotypem iiBB nebo iiB Plody dýně jsou žluté. A konečně, pokud jsou oba geny reprezentovány recesivními alelami, je plod dýně zelený. Stejný typ lze použít k dědění barvy srsti psů, ovcí a dalších zvířat.
Recesivní epistáze je detekován, když je exprese genu potlačena recesivními alelami jiného genu. Tento typ epistázy je ilustrován dědičností barvy srsti u domácích myší. Barva aguti (červenošedá) je určena dominantním genem A, jeho recesivní alela A dává černou barvu. Gen z nealelického páru V podporuje projev barvy a gen b je supresor a potlačuje účinek dominantní alely A a recesivní A. Myši s genotypem Abb fenotypově k nerozeznání od jedinců s genotypem aabb- vše bílé.
Interakce polymerních genů
Polymerismus– typ interakce nealelických genů, kdy na vývoj jednoho znaku působí několik párů nealelických genů. Takové geny se nazývají polymer(z řečtiny politika– hodně a meros- Část). Jsou označeny stejným písmenem, ale s různými indexy, které označují počet alelických párů v genotypu, které určují vývoj určitého znaku ( A 1 A 1 A 2 A 2).
Existují dvě varianty polymerace: se sčítacím účinkem genů a bez vzájemného zesílení genů.
Kumulativní účinek polymerních genů. V tomto případě závisí stupeň projevu znaku na počtu dominantních alel v genotypu jedince. Červená barva zrn pšenice je tedy určena dvěma nebo více páry genů. Každý z dominantních genů těchto alel určuje červenou barvu, zatímco recesivní geny určují bílou barvu zrn. Jedna dominantní alela produkuje zrna, která nejsou příliš barevná. Pokud jsou v genotypu přítomny dvě dominantní alely, intenzita barvy se zvyšuje. Pouze když je organismus homozygotní pro všechny páry recesivních genů, nejsou zrna zbarvena. Při křížení dihybridů tedy dochází ke štěpení v poměru 15 barevných ku jedné bílé. Ale z 15 malovaných bude mít jeden intenzivní červenou barvu, protože... obsahuje čtyři dominantní alely, čtyři budou o něco světlejší barvy, protože obsahují tři dominantní alely a jednu recesivní alelu v genotypech, šest je ještě světlejších se dvěma dominantními a dvě recesivní alely v genotypech, čtyři jsou ještě světlejší, protože mají pouze jeden dominantní a tři recesivní geny, to znamená, že skutečné rozdělení bude 1: 4: 6: 4: 1.
Podle této verze polymeru se u lidí dědí barva kůže, výška a váha. Podobný mechanismus dědičnosti je charakteristický pro mnoho kvantitativních, včetně ekonomicky cenných znaků: obsah cukru v kořenech řepy, obsah vitamínů v ovoci a zelenině, délka klasu obilovin, délka klasu, plodnost zvířat, produkce mléka hospodářských zvířat , produkce vajec kuřat atd.
Polymerní geny se navzájem neposilují. V tomto případě bude rozdělení v F 2 15:1. Plody pastýřské měšce tedy mohou být trojúhelníkové (dominantní) a oválné (recesivní). Tato vlastnost je řízena dvěma páry polymerních genů. Pokud má genotyp rostliny alespoň jednu dominantní alelu z prvního nebo druhého páru polymerních genů, pak bude tvar plodu trojúhelníkový ( A 1 A 2; A 1 a 2; a 1 A 2 ). Oválný tvar plodu budou mít pouze ty rostliny, které nemají ve svém genotypu jedinou dominantní alelu - a 1 a 1 a 2 a 2.
Akumulace určitých alel v genotypu tedy může vést ke změnám v expresi znaků.
Vícenásobné působení genů
 Gen často neovlivňuje jednu, ale řadu vlastností v organismu. Jev, kdy jeden gen může ovlivnit tvorbu několika charakteristik organismu, se nazývá pleiotropie(z řečtiny pleon– početnější a tropos- otočit se).
Gen často neovlivňuje jednu, ale řadu vlastností v organismu. Jev, kdy jeden gen může ovlivnit tvorbu několika charakteristik organismu, se nazývá pleiotropie(z řečtiny pleon– početnější a tropos- otočit se).
Existence tohoto fenoménu vůbec neodporuje klasické koncepci „jeden gen – jeden protein – jedna vlastnost“. Zjednodušeně lze vliv jednoho genu na několik znaků najednou znázornit následovně. V důsledku čtení informace z genu vzniká určitá bílkovina, která se pak může podílet na různých procesech probíhajících v těle a má tak mnohonásobné účinky. Například u hrachu závisí hnědá barva obalu semen a vývoj pigmentu v jiných částech rostliny a barva květů na jednom genu; u Drosophila gen, který způsobuje bílou barvu očí, ovlivňuje zesvětlení barvy těla a změny v některých vnitřních orgánech; U lidí gen zodpovědný za zrzavé vlasy určuje i světlejší barvu pleti a vzhled pih. Marfanův syndrom je způsoben autosomálně dominantním pleiotropním genem a projevuje se vysokým růstem, prodlužováním kostí prstů na rukou a nohou (pavoučí prsty), hypermobilitou kloubů, subluxací oční čočky a srdečními chorobami.
Toto vícenásobné působení je typické pro většinu genů. Neměli bychom si však představovat, že pleiotropní gen ovlivňuje každý znak stejně. Naprostá většina genů s různým stupněm pleiotropie se vyznačuje silnějším působením na jeden znak a mnohem slabším působením na jiný.
Genotyp jako integrální systém
Skutečnost štěpení hybridů v potomstvu umožňuje tvrdit, že genotyp je složen z jednotlivých prvků - genů, které lze dědit nezávisle ( diskrétnost genotypu). Genotyp přitom nelze považovat za prostý mechanický součet jednotlivých genů. Genotyp je systém interagujících genů. Přesněji řečeno, neinteragují samotné geny (úseky DNA), ale genové produkty vzniklé na jejich základě (RNA a poté proteiny). Proto je v některých případech působení různých genů relativně nezávislé, ale projev znaku je zpravidla výsledkem interakce produktů různých genů.
Genotyp jakéhokoli organismu je komplexní integrální systém interagujících genů. Tato integrita genotypu vznikla historicky v procesu evoluce druhu. V důsledku mutací se neustále objevují nové geny, vznikají nové chromozomy a dokonce i nové genomy. Nově vznikající geny mohou okamžitě interagovat s existujícími geny nebo modifikovat povahu jejich práce, a to i recesivní, to znamená, aniž by se projevily.
V důsledku toho se u každého druhu rostlin a živočichů genotyp projevuje jako ucelený systém, který se v daném okamžiku historicky vyvíjel.
II. Upevňování znalostí
Shrnutí konverzace při učení nového materiálu.
III. Domácí práce
Prostudujte si odstavec učebnice (vztah mezi geny a znaky, typy interakce alelických a nealelických genů, pleiotropie, genotyp jako integrální systém).
Vyřešte úlohy na konci odstavce učebnice.