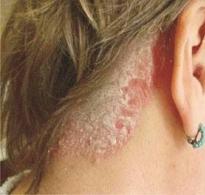
Примеры аллельных и неаллельных генов. Взаимодействие аллельных генов
Генетика человека с основами общей генетики [Учебное пособие] Курчанов Николай Анатольевич
4.3. Взаимодействие генов
4.3. Взаимодействие генов
В организме одновременно функционирует множество генов. В процессах реализации генетической информации в признак возможны многочисленные «пункты» взаимодействия разных генов на уровне биохимических реакций. Такие взаимодействия неизбежно отражаются на формировании фенотипа.
Аллельные гены определяют альтернативные признаки, так как лежат в гомологичных локусах гомологичных хромосом. Между аллелями гетерозиготы возможны определенные взаимодействия, лежащие в основе проявления признака в фенотипе. Известны три основные формы межаллельных взаимодействий.
Полное доминирование – в гетерозиготе один аллель (доминантный) подавляет проявление второго аллеля (рецессивного).
Неполное доминирование – в гетерозиготе наблюдается промежуточное проявление признаков аллелей.
Кодоминирование – независимое проявление аллелей в гетерозиготе.
В некоторых случаях механизм взаимодействия аллелей расшифрован. Лучше всего их взаимоотношения иллюстрируют белки-ферменты. Данные биохимической генетики показывают, что явление доминирования часто связано с активностью определенного фермента.
Менее понятно явление моногенного гетерозиса, когда в гетерозиготе признак проявляется сильнее, чем в гомозиготе по доминантному аллелю. Гетерозис – явление превосходства гибридов над обеими родительскими формами. Феномен гетерозиса интенсивно изучался в связи с его важной ролью для селекции. Это явление весьма сложное и неоднозначное. Неоднозначно понятие «превосходство», так как гетерозис проявляется на репродуктивном, соматическом и адаптивном уровнях. Проявления на разных уровнях могут быть противоположной направленности (например, «улучшение» соматических показателей может сопровождаться «ухудшением» адаптивных). Неоднозначны сами понятия «улучшение», «ухудшение», поскольку они не имеют четких критериев и их применение крайне субъективно. Существует несколько теорий, объясняющих природу гетерозиса.
Феномен моногенного гетерозиса, когда показана зависимость признака от одной аллельной пары, является только одним аспектом теории гетерозиса. По предложению Ф. Добжанского это явление получило название сверхдоминирования. Оно имеет большое значение для эволюционной теории, поскольку демонстрирует преимущество гетерозигот в популяциях. Однако это понятие скорее применимо к адаптивному уровню, поэтому до выяснения генетических механизмов сверхдоминирования рассматривать его как особый вид межаллельных взаимодействий преждевременно.
В живых организмах часто взаимодействуют не только аллели одного гена, но и аллели разных генов, давая самые различные варианты расщепления. Различают три основных типа взаимодействия неаллельных генов.
Комплементарность – взаимодействие разных доминантных аллелей обусловливает появление нового признака.
По типу комплементарности обычно взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Однако для некоторых морфологических признаков биохимический механизм реализации неизвестен.
Эпистаз – один ген подавляет проявление другого, неаллельного ему гена.
Гены, подавляющие действие других генов, называются эпистатическими (или генами-супрессорами). Возможны два варианта эпистаза: доминантный эпистаз – эпистатический ген является доминантным в своей аллельной паре и рецессивный эпистаз – эпистатический ген является рецессивным в своей аллельной паре.
Полимерия – однозначное действие неаллельных генов. Полимерия связана с контролем признака несколькими неаллельными генами. Полигенный контроль весьма широко распространен в генетике. Полимерные гены обычно обозначаются одинаковыми буквами с нижним индексом – А 1 , А 2, А 3 и т. д.
Полимерия также встречается в двух вариантах. При кумулятивной полимерии интенсивность признака пропорциональна числу доминантных аллелей среди полимерных генов, а при некумулятивной полимерии разные полимерные гены дублируют друг друга и для проявления признака достаточно наличия одного из доминантных аллелей.
Многочисленные случаи взаимодействия генов заполняют основной объем всех задачников по генетике. В типичном случае при скрещивании дигетерозигот при взаимодействии генов образуются самые различные отношения фенотипических классов в поколениях – 9: 3: 4; 9: 7; 13: 3; 12: 3: 1; 15: 1 и другие. Генетический анализ показывает, что все они являются видоизменением классической менделевской формулы дигибридного расщепления 9: 3: 3: 1. Решение большого количества задач по генетике является необходимым этапом в подготовке студентов, изучающих генетику.
Словосочетание «взаимодействие генов» несколько условно, так как обычно взаимодействуют не сами гены, а их продукты. Однако нельзя согласиться с термином «взаимодействие фенов», который неточно отражает смысл явления. На мой взгляд, в учебной литературе лучше оставить традиционный термин «взаимодействие генов» (аллельных и неаллельных).
Данный текст является ознакомительным фрагментом. Из книги Генетика окрасов собак автора Робинсон РойВзаимодействие локусов А и Е Важность этих двух серий аллелей будет показана в данном разделе. Обе серии контролируют распределение черного и желтого пигментов по шерсти у большинства пород собак. Во взаимодействии эти локусы образуют множество хорошо известных
Из книги Микробиология: конспект лекций автора Ткаченко Ксения Викторовна2. Взаимодействие вирусов с клеткой хозяина Взаимодействие идет в единой биологической системе на генетическом уровне.Существует четыре типа взаимодействия:1) продуктивная вирусная инфекция (взаимодействие, в результате которого происходит репродукция вируса, а
Из книги Муравей, семья, колония автора Захаров Анатолий АлександровичВЗАИМОДЕЙСТВИЕ МУРАВЬЕВ В СЕМЬЕ Семья муравьев - объединение сотен, тысяч, а иногда и миллионов индивидов, усилиями которых сооружается гнездо, выкармливается многочисленное потомство, охраняется муравейник и его кормовой участок от посягательств агрессивных
Из книги Эволюция автора Дженкинс МортонДРЕЙФ ГЕНОВ Иногда эта концепция называется «эффект Сьюэлла - Райта», в честь предложивших ее двух популяционных генетиков. После того как Мендель доказал, что гены являются единицами наследственности, а Харди и Вайнберг продемонстрировали механизм их поведения,
Из книги Род человеческий автора Барнетт ЭнтониВзаимодействие наследственности и среды Иногда спрашивают: что важнее - наследственность или окружающая среда? На этот вопрос не так легко ответить. Если под этим подразумевать, чт? имеет наибольшую силу воздействия, то и тогда следует ограничиться частными случаями.
Из книги Основы психофизиологии автора Александров Юрий7. ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровне. Особенно широка интеграция сигналов в ретикулярной формации. В коре мозга происходит интеграция сигналов высшего порядка. В
Из книги Эмбрионы, гены и эволюция автора Рэфф Рудольф АВзаимодействие и интеграция В наши дни посетители музеев воспринимают выставленные в них смонтированные скелеты огромных вымерших животных как нечто само собой разумеющееся. Однако так было не всегда. Научные сведения о таких импозантных формах, как мамонты и
Из книги Теория адекватного питания и трофология [таблицы текстом] автора Из книги Стой, кто ведет? [Биология поведения человека и других зверей] автора Жуков. Дмитрий АнатольевичВзаимодействие эндокринных систем: прямая связь, обратная связь, синергизм, пермиссивное действие, антагонизм Как уже было упомянуто, отдельные эндокринные железы и их гормоны взаимодействуют друг с другом, обеспечивая функциональное единство организма. Между
Из книги Теория адекватного питания и трофология [таблицы картинками] автора Уголев Александр Михайлович9.7. Заключительные замечания (взаимодействие в биоценозах) Рассмотрим биохимические взаимодействия живых организмов в естественных условиях, которые распространены чрезвычайно широко и стали объектом специальной науки - аллелопатии (см. гл. 1). Примеры таких
Из книги Фенетика [Эволюция, популяция, признак] автора Яблоков Алексей ВладимировичВзаимодействие эволюционных сил - механизм эволюции Пусковой механизм эволюции функционирует в результате совместного действия эволюционных факторов в пределах популяции как эволюционной единицы. Всякая популяция у любого вида подвержена тому или иному давлению
Из книги Эволюция [Классические идеи в свете новых открытий] автораДупликация генов МНОГОФУНКЦИОНАЛЬНЫЕ ГЕНЫ - ОСНОВА ЭВОЛЮЦИОННЫХ НОВШЕСТВ.Мысль о том, что дупликация генов служит важнейшим источником эволюционных новшеств, была высказана еще в 1930-е годы выдающимся биологом Джоном Холдейном (Haldane, 1933). Сегодня в этом нет никаких
Из книги Гены и развитие организма автора Нейфах Александр Александрович1. Промоторы генов В этом разделе мы кратко расскажем о том, какие нуклеотидные последовательности, прилегающие к генам, а иногда и внутри гена, ответственны за процесс транскрипции. У прокариот эти участки, с которыми связывается молекула РНК-полимеразы и откуда
Из книги Эволюция человека. Книга 1. Обезьяны, кости и гены автора Марков Александр ВладимировичИзменения активности генов Эволюция животных в целом и приматов в частности протекает не столько за счет изменения структуры белок-кодирующих генов, сколько за счет изменения их активности. Небольшое изменение в верхних этажах иерархически организованных
Из книги Генетика человека с основами общей генетики [Учебное пособие] автора Курчанов Николай Анатольевич4.4. Взаимодействие генотипа и среды Природа проявления действия генов намного сложнее, чем в описанных выше вариантах. Рассматривая действие генов и их аллелей, необходимо учитывать влияние внешней среды на проявление признаков, а также модифицирующее действие других
Из книги автора7.1. Выделение генов Возможно использование нескольких путей выделения генов. Каждый из них имеет свои достоинства и недостатки.Химический синтез генов, т. е. синтез нуклеотидов с заданной последовательностью, соответствующей одному гену, впервые был осуществлен в
Взаимодействие между аллельными генами осуществляется в виде трех форм: полное доминирование, неполное доминирование и независимое проявление (кодоминирование).
Полное доминирование – когда один доминантный аллель полностью подавляет проявление рецессивного аллеля, например, желтая окраска горошин доминирует над зеленой.
Неполное доминирование наблюдается в том случае, когда один ген из пары аллелей не обеспечивает образование в достаточном для нормального проявления признака его белкового продукта. При этой форме взаимодействия генов все гетерозиготы и гомозиготы значительно отличаются по фенотипу друг от друга. Примером расщепления при неполном доминировании может служить наследование окраски цветков Ночной красавицы.
При скрещивании растений с красными цветками (АА) и растений с белыми (аа) гибриды F1 имеют розовые цветки (Аа). Таким образом, имеет место неполное доминирование; в F2 наблюдается расщепление 1: 2: 1 как по фенотипу, так и по генотипу.
Кроме полного и неполного доминирования известны случаи отсутствия доминантно-рецессивных отношений или кодоминирования. При кодоминировании у гетерозиготных организмов каждый из аллельных генов вызывает формирование в фенотипе контролируемого им признака.
Примером этой формы взаимодействия аллелей служит наследование групп крови человека по системе АВ0, детерминируемых геном I. Существует три аллеля этого гена Io, Ia, Ib, определяющие антигены групп крови. Наследование групп крови иллюстрирует также явление множественного аллелизма: в генофондах популяций человека ген I существует в виде трех разных аллелей, которые комбинируются у отдельных индивидуумов только попарно. До этого примера мы говорили о генах, существующих только в двух разных аллельных формах. Однако многие гены состоят из сотен пар нуклеотидов, так что мутации могут проходить во многих участках гена и порождать множество различных его аллельных форм. Так как в каждой из гомологичной хромосом имеется по одному аллельному гену, то, разумеется, диплоидный организм имеет не более двух из серии аллелей генофонда популяции.
30. Неаллельные взаимодействия генов
Неаллельные гены - это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов:
комплемментарность;
полимерия.
Комплементарное (дополнительное) действие генов - это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных - удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 - сферические и 1 - удлинённые.
Эпистаз - взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый - гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз - это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3.
Полимерия - взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 но фенотипу происходит в соотношении 1:4:6:4:1.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1.
Пример: цвет кожи у людей, который зависит от четырёх генов.
Аллельные гены – гены, расположенные в одинаковых участках гомологичных хромосом и контролирующие развитие вариаций одного признака.
Неаллельные гены – расположены в разных участках гомологичных хромосом, контролируют развитие разных признаков.
Понятие о действии генов.
Ген – участок молекулы ДНК или РНК, кодирующий последовательность нуклеотидов в тРНК и рРНК или последовательность аминокислот в полипептиде.
Характеристики действия генов:
Ген дискретен
Ген специфичен – каждый ген отвечает за синтез строго определенного вещества
Ген действует градуально
Плейотропное действие – 1 ген действует на изменение или проявление нескольких признаков (1910 Плате) фенилкетонурия, синдром Марфана
Полимерное действие – для экспрессивности признака нужно несколько генов (1908 Нильсон-Эле)
Гены взаимодействуют между собой через белковые продукты, детерминированные ими
На проявление генов оказывают влияние факторы среды
Перечислите типы взаимодействия между аллельными и неаллельными генами.
Между аллельными:
Полное доминирование
Неполное доминирование
Кодоминирование
Сверхдоминирование
Между неаллельными: (признак или свойства детерминируются двумя или более неаллельными генами, которые взаимодействуют между собой. Хотя и здесь взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими продукты. При этом имеет место отклонение от менделеевских закономерностей расщепления).
Полимерия
Комплиментарность
Сущность полного доминирования. Примеры.
Полное доминирование – тип взаимодействия аллельных генов, при котором доминантный ген (А) полностью подавляет действие рецессивного гена (а) (веснушки)
Неполное доминирование. Примеры.
Неполное доминирование – тип взаимодействия аллельных генов, при котором доминантный аллель не полностью подавляет действие рецессивного аллеля, формируя признак с промежуточной степенью вырожденности (цвет глаз, форма волос)
Сверхдомининрование как основа гетерозиса. Примеры.
Сверхдоминирование – тип взаимодействия аллельных генов, при котором ген, находящийся в гетерозиготном состоянии имеет большее фенотипическое проявление признака, чем гомозиготный.
Серповидно-клеточная анемия. А – гемоглобинA, а – гемоглобинS. АА – 100% нормальные эритроциты, больше подвержены малярии; аа – 100% мутированные (умирают), Аа – 50% мутированных, практически не подвержены малярии т.к. уже поражены
Кодоминирование и его сущность. Примеры.
Кодоминирование – тип взаимодействия аллельных генов, при котором в детерминации признака участвуют несколько аллелей гена и происходит формирование нового признака. Один аллельный ген дополняет действие другого аллельного гена, новый признак отличается от родительских (группы крови АВО).
Явление независимого друг от друга проявления обоих аллелей в фенотипе гетерозиготы, иными словами - отсутствие доминантно-рецессивных отношений между аллелями. Наиболее известный пример - взаимодействие аллелей, определяющих четвертую группу крови человека (АВ). Известна множественная серия, состоящая из трех аллелей гена I, определяющего признак группы крови человека. Ген I отвечает за синтез ферментов, присоединяющих к белкам, находящимся на поверхности эритроцитов, определенные полисахариды. (Этими полисахаридами на поверхности эритроцитов как раз и определяется специфичность групп крови.) Аллели 1 А и 1 в кодируют два разных фермента; аллель 1° не кодирует никакого. При этом аллель 1° рецессивен и по отношению к 1 А, и по отношению к I B , а между двумя последними нет доминантно-рецессивных отношений. Люди, имеющие четвертую группу крови, несут в своем генотипе два аллеля: 1 А и 1 B . Поскольку между этими двумя аллелями нет доминантно-рецессивных отношений, то в организме таких людей синтезируются оба фермента и формируется соответствующий фенотип - четвертая группа крови.
Полное доминирование.
На примере опытов Г. Менделя с горохом видно, что один ген может полностью подавлять проявление другого аллельного гена.
Неполное доминирование.
При неполном доминировании признак и первом поколении проявляется промежуточно между родительскими формами. Если спаривать длинноухих каракульских овец с безухими, то в F1 все потомки будут короткоухими. Расщепление в F2 по фенотипу и генотипу совпадает (1:2:1).
Кодоминирование.
В фенотипе проявляются оба аллельных гена, не обнаруживая между собой ни доминантности, ни рецессивности. У человека известны системы крови АВО, которые обусловливают эритроцитарные антигены. Аллели А и В взаимодействуют по типу кодоминирования.
С 1940 но 1970 г. у крупного рогатого скота открыто 12 генетических систем групп крови. В них различают более 100 антигенов крови, определяющих 369 феногрупп, которые могут составлять около двух триллионов различных сочетаний - серологических типов. Феногруппа
- группа антигенов (иногда один антиген), которые являются фенотипическим выражением совокупности отдельных генов одного локуса. Группы крови, в основе которых лежат индивидуальные особенности антигенных свойств эритроцитов, наследственно обусловлены и не меняются в течение всей жизни животного. Наследуются они одиночно или комплексно и поэтому могут служить удобной генетической моделью в решении многих теоретических и практических вопросов селекции, так как подавляющее количество известных аллельных эритоцитарных факторов наследуются по типу кодоминирования.
Сверхдоминирование.
Оно характеризуется тем, что у гетерозиготных особей признак проявляется сильнее, чем у гомозиготных доминантных, то есть гибрид превосходит обоих родителей по развитию признака. Такое явление называется гетерозасом (Аа>АА и а а). Наглядный пример сверхдоминирования - серповидною[с точная анемия человека. Гомозиготы SS имеют часть эритроцитов в форме серпа и стрелолиста, у них нарушена транспортировка кислорода эритроцитами. Люди страдают острой формой анемии и обычно умирают в раннем возрасте. Гетерозиготы Ss устойчивы к тропической малярии, гомозиготы ss заболевают тропической малярией.
Множественный аллелизм.
Мы рассматривали в основном взаимодействие двух аллельных генов, из которых каждый может изменяться в результате мутации и возникать новый аллельный ген. Это явление получило название "множественный аллелизм". У человека примером множественного аллелизма служит система крови АВО, имеющая 3 аллельных гена (А, В, О) и соответствующим образом 6 генотипов и 4 фенотипа.
У кроликов установлены следующие аллели, влияющие на окраску волосяного покрова: С - черный, ссh - шиншилла, сh - гималайский, са - альбинос. Аллели представлены в порядке убывания доминантности (С>cch>ch>сa). Аллель С доминирует над всеми последующими, сa - рецессивен ко всем предыдущим (рис. 2.5).
Плейотропное действие гена. Ген может влиять на развитие двух и более признаков, то есть наблюдается множественное действие гена. Такой тип взаимодействия был обнаружен при разведении каракульских овец серой окраски (ширази) и черной (араби). Ягнята, гомозиготные по доминантному аллелю гена С, определяющему серую окраску (ширази). при переходе на подножный корм погибают из-за недоразвития парасимпатической нервной системы рубца. Это обусловлено действием двух доминантных генов, которые обладают двойным (плейотропным) действием: обусловливают серую окраску и в гомозиготном состоянии - дефект нервной системы.
Академик Д. К. Беляев с сотрудниками установил, что у норок некоторые мутации, связанные с изменением окраски волосяного покрова, рецессивны, и в силу плейотропного действия при этом снижаются плодовитость и жизнеспособность животных.
Летальные и полулегальные гены. Гены вызывают сложные физиологические изменения в процессе развития организма. Они могут понижать жизнеспособность организма вплоть до летального исхода.
Одной из причин, изменяющих расщепление в соотношении 3:1. является разная жизнеспособность гомозигот в F2. В XX в. были обнаружены факты, когда в происходило расщепление по фенотипу 2:1. Так, при изучении наследования окраски у желтых и черных мышей наблюдали расщепление по окраске на 2 части желтых и 1 часть черных мышей. При этом часть желтых мышей погибала в эмбриональный период. Один и тот же ген в доминантном гетерозиготном состоянии обусловливал желтую окраску мышей, а в гомозиготном состоянии проявлял свое летальное действие. Ген черной окраски является аллельным и рецессивным.
В 30-х годах XX в. была обнаружена новая окраска у серебристо-черной лисицы, названная платиновой. При скрещивании платиновых лисиц между собой особи имели как платиновую, так и серебристо-черную окраску в соотношении 2:1. При проведении анализирующего скрещивания стало ясно, что ген платиновой окраски является доминантным. Было высказано предположение, что гомозиготные платиновые лисицы (АА) гибнут в эмбриональный период. Вскрытие беременных самок подтвердило это предположение.
Рецессивные летальные гены действуют в гомозиготном состоянии, а доминантные - и в гетерозиготном. Если ген не вызывает гибели организма, но значительно снижает жизнеспособность, то его называют полулетальным.
Летальные и полулегальные гены чаше всего рецессивные. Летальные гены вызывают гибель организма в эмбриональный или постэмбриональный период. Например, рецессивные легальные гены обусловливают отсутствие шерстного покрова у новорожденных телят, крольчат и животных других видов. Вскоре после рождения эти животные погибают.
Полулегальные гены могут вызвать уродства у животных: укорочение осевого скелета; деформацию головы, конечностей; нарушение физиологических функции организма. Племенных животных, имеющих летальные и полулетальные гены, необходимо выбраковывать. Особенно тщательно необходимо проверять племенных производителей, от которых при искусственном осеменении получают большое число потомков.
Легальные гены могут беспрепятственно распространяться из одной страны в другую. Так, племенного быка голландской породы Принца Адольфа, носителя генетической аномалии, завезли в Швецию из Нидерландов в 1902 г. Спустя 26 лет стало известно, что бык является носителем летального рецессивною гена бесшерстности телят. Его дочерей, сыновей и внуков широко использовали для племенных целей, и через относительно короткое время многие животные были родственны Принцу Адольфу. В некоторых стадах около 6 % телят рождались без шерсти и через несколько минут после рождения погибали.
В последнее время в зарубежной и отечественной литературе появились сообщения о генетических аномалиях (дефектах), наносящих огромный экономический ущерб племенному животноводству (P. D. Smith, Л. Ф. Новикова). Гак, например, дефекты ВLAD (Bovine Leukocyte Adhesion Deficiency), или адгезия (слипание) лейкоцитов у крупного рогатого скота и CVM (Complex Vertebral Malformation), или комплексное уродство позвоночника имеют рецессивный тип наследования и не выражены в гетерезиготном состоянии (рис. 2.6). Значительное распространение данные генетические дефекты получили в линии голштинского скота Монтвик Чифтейн 95679, которую широко используют в Российской Федерации. Один из самых известных быков этой линии Осбордейл Айвенго 1189870 являлся носителем генетического дефекта В LAD. a егo внук Карлин-М Айвенго Бэлл 1667366 - носителем обоих генетических дефектов BLAD и CVM.
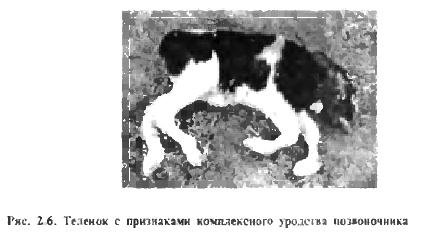
В среднем 2-5 % новорожденных животных многих видов имеют наследственные аномалии. Летальные и полулегальные гены - одна из причин эмбриональной смертности и врожденных аномалий. Выявлено несколько десятков таких генов по каждому виду сельскохозяйственных животных, дана международная классификация аномалий. О наследственной обусловленности мертворождений, выкидышей, уродств говорит тот факт, что их процент значительно повышается при инбридинге.
Наиболее распространенные наследственные аномалии у крупного рогатого скота следующие: мопсовидность; контрактура мышц; отсутствие передних и задних конечностей; водянка плода и головного мозга; дефекты кожного покрова: бесшерстность; укорочение челюсти; пупочная грыжа и т. д.
Обобщая данные разных авторов, можно сказать, что у крупного рогатого скота имеют место более 40 гомозиготных леталей, уродств и аномалий кариотипа - наследственного брака. Выявлено 23 врожденных дефекта, влияющих на воспроизводительную функцию и плодовитость коров.
У свиней описано 33 генетические аномалии. Наиболее часто встречаются: мозговая грыжа; паралич задних конечностей; отсутствие анального отверстия; волчья пасть; толстоногость; искривление и ригидность конечностей; водянка головного мозга; гемофилия.
У овец наиболее выражены черепно-лицевые дефекты: укорочение или отсутствие нижней челюсти; выпадение шерсти, облысение; карликовость; волчья пасть и др. Всего 43 аномалии.
В Международный список из легальных дефектов включено 45 аномалий у кур; 6 - у индеек; 3 - у уток; из наследственных аномалий 10 -у лошадей.
На наследственное происхождение аномалии указывают следующие факты: соответствие обнаруженной аномалии описанию генетического дефекта, появление аномалии в результате инбридинга, проявление подобных аномалий у боковых родственников (сибсов и полусибсов), родственников матери или отца.
Все случаи проявления наследственных аномалий необходимо фиксировать в карточках племенных животных, чтобы в дальнейшем можно было провести генетический анализ. В данной ситуации большую роль играют родословная и объективный племенной учет Сбор данных для опенки генетическою благополучия племенных животных и наличия летальных и полулегальных генов следует проводить на племенных предприятиях, станциях по искусственному осеменению сельскохозяйственных животных.
Взаимодействие аллельных генов и его типы
Изучая законы Г.Менделя, мы уже знакомились с основными типами взаимодействия аллельных генов. На основе ранее изученного материала заполните таблицу.
Таблица. Типы взаимодействия генов одной аллельной пары
Тип взаимодействия |
Характер взаимодействия в |
Формулы расщепления по фенотипу в F 2 и при анализирующем скрещивании |
Примеры |
| Полное доминирование | Один аллель (доминантный) полностью подавляет действие другого (рецессивного) аллеля |
Высокий рост растения доминирует над карликовостью; вьющиеся волосы – над прямыми; карие глаза – над голубыми и т. п. |
|
| Неполное доминирование | Действие каждого аллеля реализуется не полностью, вследствие чего проявляется промежуточный признак |
Окраска цветков ночной красавицы; серповидноклеточная анемия; цистонурия и т.п. |
|
| Кодоминирование | Участие обоих аллелей в определении признака |
IV группа крови (эритроциты этой группы крови имеют агглютиногены типа А и типа В ) |
Взаимодействие неаллельных генов
Исходя из законов Г.Менделя, можно прийти к заключению, что существует довольно прочная связь между геном и признаком, что генотип слагается из суммы независимо действующих генов, а фенотип – механическая совокупность отдельных признаков. Однако прямые и однозначные связи гена с признаком скорее исключение, чем правило. Развитие признака организма обычно находится под контролем многих генов, и признак является результатом взаимодействия неаллельных генов.
Простейший случай взаимодействия неаллельных генов – когда признак контролируется двумя парами аллелей. Рассмотрим некоторые примеры такого взаимодействия.
Комплементарное взаимодействие генов
Явление взаимодействия генов, при котором для проявления признака необходимо наличие двух неаллельных генов, называют комплементарностью (от лат. complementum – дополнение), а гены, необходимые для проявления признака, комплементарными , или дополнительными.
Рассмотрим различные варианты комплементарного взаимодействия генов.
Аллели второго гена проявляются только при наличии доминантного аллеля гена А . Окраска мышей зависит в простейшем случае от двух генов. Мыши с генотипом аа лишены пигмента и имеют белый цвет. При наличии доминантного аллеля А пигмент вырабатывается, и мышь как-то окрашена. Конкретный цвет определяется вторым геном. Его доминантный аллель С обусловливает серый цвет мыши, а рецессивный аллель с – черный цвет. Таким образом, если рассматривать гомозиготные варианты, генотип серых мышей – ААСС , черных – ААсс , белых – ааСС или аасс . При скрещивании серой мыши ААСС с белой аасс в первом поколении все получается по Менделю: все гибриды имеют серый цвет (это гетерозиготы с генотипом АаСс ). В F2, как легко проверить, получим 9/16 серых мышей, 3/16 черных и 4/16 белых.

Для проявления признака в генотипе должны присутствовать доминантные аллели двух разных генов. Примером такого комплементарного взаимодействия генов является наследование формы плода у тыквы. При наличии обоих доминантных аллелей плоды имеют дисковидную форму, при наличии одного (любого!) доминантного аллеля – сферическую, а при отсутствии доминантных аллелей – удлиненную.

Для проявления признака в генотипе должны присутствовать доминантные аллели двух разных генов, но каждый доминантный аллель в сочетании с рецессивными аллелями другой пары имеет самостоятельное фенотипическое проявление. Например, у кур гороховидная форма гребня определяется одним доминантным геном, розовидная – другим неаллельным ему, но тоже доминантным геном. Когда эти гены окажутся в одном генотипе, развивается ореховидная форма гребня. В случае если организм оказывается гомозиготным по обоим рецессивным генам, развивается простой листовидный гребень. При скрещивании дигибридов (все с ореховидным гребнем) во втором поколении происходит расщепление в отношении 9:3:3:1. Но здесь нельзя найти независимого расщепления каждого аллеля в отношении 3:1, так как во всех случаях совпадения в генотипе обоих доминантных генов их прямого действия не обнаруживается.
Таким образом, о комплементарном взаимодействии неаллельных генов говорят в том случае, когда два неаллельных гена дают новый признак, то есть имеет место новообразование.
Эпистатическое взаимодействие генов
Взаимодействие генов, при котором один ген подавляет действие другого, неаллельного первому, называется эпистазом (от греч. эпи – над и стазис – стоять), а гены, подавляющие действие других генов, называются генами-супрессорами , или генами-ингибиторами (от лат. inhiber – удерживать), или эпистатическими генами . Подавляемый ген называется гипостатическим .
Различают эпистаз доминантный и рецессивный.
Доминантный эпистаз связан с доминантным геном-ингибитором. Например, ген I обусловливает белый цвет плодов тыквы, при его наличии действие гена В не проявляется. При генотипе iiВВ или iiВb плоды тыквы имеют желтый цвет. Наконец, если оба гена представлены рецессивными аллелями, плоды тыквы имеют зеленый цвет. По этому же типу может наследоваться окраска шерсти собак, овец и других животных.
Рецессивный эпистаз обнаруживается тогда, когда проявление гена подавляется рецессивными аллелями другого гена. Этот вид эпистаза иллюстрируется наследованием окраски шерсти у домовых мышей. Окраска агути (рыжевато-серая) определяется доминантным геном А , его рецессивный аллель а дает черную окраску. Ген из неаллельной пары В способствует проявлению цветности, а ген b является супрессором и подавляет действие доминантного аллеля А и рецессивного а . Мыши с генотипом Аbb неотличимы по фенотипу от особей с генотипом ааbb – все белые.
Полимерное взаимодействие генов
Полимерия – тип взаимодействия неаллельных генов, при котором несколько пар неаллельных генов действуют на развитие одного признака. Такие гены называются полимерными (от греч. полис – много и мерос – часть). Их обозначают одной буквой, но с разными индексами, которые указывают на число аллельных пар в генотипе, обусловливающих развитие конкретного признака (А 1 А 1 А 2 А 2 ).
Различают два варианта полимерии: с суммирующим действием генов и без усиления генов друг другом.
Суммирующее действие полимерных генов. В этом случае степень проявления признака зависит от числа доминантных аллелей в генотипе особи. Так, красная окраска зерен пшеницы определяется двумя и более парами генов. Каждый из доминантных генов этих аллелей определяет красную окраску, рецессивные гены определяют белый цвет зерен. Один доминантный аллель дает не очень сильно окрашенные зерна. Если в генотипе присутствуют два доминантных аллеля, интенсивность окраски повышается. Лишь в том случае, когда организм оказывается гомозиготным по всем парам рецессивных генов, зерна не окрашены. Таким образом, при скрещивании дигибридов происходит расщепление в отношении 15 окрашенных к одному белому. Но из 15 окрашенных один будет иметь интенсивный красный цвет, т.к. содержит четыре доминантных аллеля, четыре будут окрашены несколько светлее, т.к. содержат в генотипах три доминантных аллеля и один рецессивный аллель, шесть – еще светлее с двумя доминантными и двумя рецессивными аллелями в генотипах, четыре – еще более светлые, т.к. имеют лишь один доминантный и три рецессивных гена, то есть истинное расщепление будет 1:4:6:4:1.
По данному варианту полимерии наследуются окраска кожи, рост и масса у человека. Подобный же механизм наследования характерен для многих количественных, в том числе и хозяйственно-ценных признаков: содержание сахара в корнеплодах свеклы, содержание витаминов в плодах и овощах, длина колоса злаков, длина початка кукурузы, плодовитость животных, молочность скота, яйценоскость кур и др.
Полимерные гены не усиливают друг друга. В этом случае расщепление в F 2 будет 15:1. Так, плоды пастушьей сумки могут быть треугольными (доминантный признак) и овальными (рецессив). Признак контролируется двумя парами полимерных генов. Если в генотипе растения имеется хотя бы один доминантный аллель из первой или второй пары полимерных генов, то форма плода у него будет треугольной (А 1 А 2 ; А 1 а 2 ; а 1 А 2 ). Овальную форму плодов будут иметь лишь те растения, у которых в генотипе нет ни одного доминантного аллеля – а 1 а 1 а 2 а 2 .
Таким образом, накопление определенных аллелей в генотипе может привести к изменению выраженности признаков.
Множественное действие генов
 Часто ген оказывает действие не на один, а на ряд признаков организма. Явление, при котором один ген может влиять на формирование нескольких признаков организма, называется плейотропией
(от греч. плеон
– более многочисленный и тропос
– поворот).
Часто ген оказывает действие не на один, а на ряд признаков организма. Явление, при котором один ген может влиять на формирование нескольких признаков организма, называется плейотропией
(от греч. плеон
– более многочисленный и тропос
– поворот).
Существование этого явления отнюдь не противоречит классической концепции «один ген – один белок – один признак». Упрощенно влияние одного гена сразу на несколько признаков можно представить следующим образом. В результате считывания информации с гена образуется некий белок, который затем может участвовать в различных процессах, происходящих в организме, оказывая таким образом множественное действие. Например, у гороха бурая окраска кожуры семян и развитие пигмента в других частях растения, окраска цветков зависят от одного гена; у дрозофилы ген, обусловливающий белую окраску глаз, влияет на осветление окраски тела и изменение некоторых внутренних органов; у человека ген, отвечающий за рыжий цвет волос, одновременно определяет более светлую окраску кожи и появление веснушек. Синдром Марфана обусловлен аутосомным доминантным плейотропным геном и проявляется высоким ростом, удлинением костей пальцев рук и ног (паучьи пальцы), гиперподвижностью суставов, подвывихом хрусталика глаза, пороком сердца.
Такое множественное действие характерно для большинства генов. Однако не следует представлять, что плейотропный ген в равной степени влияет на каждый из признаков. Для абсолютного большинства генов с той или иной степенью плейотропии характерно более сильное влияние на один признак и значительно более слабое – на другой.
Генотип как целостная система
Факт расщепления в потомстве гибридов позволяет утверждать, что генотип слагается из отдельных элементов – генов, которые могут наследоваться независимо (дискретность генотипа ). В то же время генотип не может рассматриваться как простая механическая сумма отдельных генов. Генотип – это система взаимодействующих генов. Точнее, взаимодействуют не сами гены (участки ДНК), а образуемые на их основе генные продукты (РНК, а затем – белки). Поэтому в отдельных случаях действие разных генов относительно независимо, но, как правило, проявление признака есть результат взаимодействия продуктов разных генов.
Генотип любого организма представляет собой сложную целостную систему взаимодействующих генов. Эта целостность генотипа возникла исторически в процессе эволюции вида. В результате мутаций постоянно появляются новые гены, формируются новые хромосомы и даже новые геномы. Вновь возникшие гены могут сразу же взаимодействовать с уже имеющимися генами или модифицировать характер работы последних, даже будучи рецессивными, то есть, не проявляясь сами по себе.
Следовательно, у каждого вида растений и животных генотип проявляет себя как исторически сложившаяся к данному моменту целостная система.
II. Закрепление знаний
Обобщающая беседа по ходу изучения нового материала.
III. Домашнее задание
Изучить параграф учебника (связь между генами и признаками, типы взаимодействия аллельных и неаллельных генов, плейотропия, генотип как целостная система).
Решить задачи в конце параграфа учебника.